Vous aimerez peut-être aussi
- Inmunidad Innata Elementos Constitutivos y Demas PDFDocument41 pagesInmunidad Innata Elementos Constitutivos y Demas PDFlordgimli7767% (3)
- Clasificación PRRDocument3 pagesClasificación PRRFernando Sanchez MorenoPas encore d'évaluation
- Sangre: Composición, Grupos Sanguíneos y Pruebas TransfusionalesDocument80 pagesSangre: Composición, Grupos Sanguíneos y Pruebas TransfusionalesMaribel Angela Palomino Patilla100% (2)
- Mecanismos de Defensa Del Huésped Frente A La InfecciónDocument2 pagesMecanismos de Defensa Del Huésped Frente A La InfeccióngsaPas encore d'évaluation
- Metabolismo CerebralDocument12 pagesMetabolismo CerebralFranklinYanes100% (3)
- RinovirusDocument13 pagesRinovirusMichelleMaciasPas encore d'évaluation
- InmunoglobulinasDocument20 pagesInmunoglobulinasAndrea Del Rosario MaldonadoPas encore d'évaluation
- INFLAMACION y FAGOCITOSISDocument7 pagesINFLAMACION y FAGOCITOSISYoly Vasquez100% (2)
- Citocinas PDFDocument5 pagesCitocinas PDFCris Dev100% (1)
- Mecanismo de Evasión Por HongosDocument2 pagesMecanismo de Evasión Por HongosFabrizio Falcon CeferinoPas encore d'évaluation
- CromoblastomicosisDocument11 pagesCromoblastomicosisJosé Zarur BartoloméPas encore d'évaluation
- Linfocitos T TB PDFDocument61 pagesLinfocitos T TB PDFingrid QuiñonesPas encore d'évaluation
- Procesamiento Presentacion Antigenica PDFDocument28 pagesProcesamiento Presentacion Antigenica PDFRichard Bravo CéspedesPas encore d'évaluation
- Inflamación GranulomatosaDocument6 pagesInflamación GranulomatosaperezdcPas encore d'évaluation
- Inflamación y Reparación TisularDocument11 pagesInflamación y Reparación TisularBetty CarreñoPas encore d'évaluation
- MACROFAGOSDocument6 pagesMACROFAGOSCarlitos AyalaPas encore d'évaluation
- Activación de Los Linfocitos TDocument11 pagesActivación de Los Linfocitos TmariaPas encore d'évaluation
- Presentación antigénica y moléculas MHCDocument44 pagesPresentación antigénica y moléculas MHCEsmeralda RVPas encore d'évaluation
- Casos Clínicos VirologíaDocument8 pagesCasos Clínicos VirologíaARELY MURIEL MORENOPas encore d'évaluation
- Acumulaciones IntracelularesDocument10 pagesAcumulaciones IntracelularesJR Bio GenPas encore d'évaluation
- Opsonización PDFDocument4 pagesOpsonización PDFSelena Cedeño PicoPas encore d'évaluation
- Neisseria meningitidis: morfología, diagnóstico, transmisión, manifestaciones y prevenciónDocument3 pagesNeisseria meningitidis: morfología, diagnóstico, transmisión, manifestaciones y prevenciónYaritza PinargtPas encore d'évaluation
- StreptococcusDocument21 pagesStreptococcus.:("*"BLacK BuLLeT"*"):.100% (5)
- Anemia Hemolítica Autoinmunitaria Como Manifestación Inicial de Artritis Reumatoide. Reporte de Un CasoDocument5 pagesAnemia Hemolítica Autoinmunitaria Como Manifestación Inicial de Artritis Reumatoide. Reporte de Un CasoWer Yusheim ShinigamiPas encore d'évaluation
- INMUNIDAD INNATA: DEFENSA PRIMARIA CONTRA MICROBIOSDocument56 pagesINMUNIDAD INNATA: DEFENSA PRIMARIA CONTRA MICROBIOSMelody Maza67% (3)
- Bacterias ResumenDocument74 pagesBacterias ResumenRamón H. JuárezPas encore d'évaluation
- Fagocitosis ExpoDocument15 pagesFagocitosis ExpoVíctor SaldañaPas encore d'évaluation
- Estructura BacterianaDocument14 pagesEstructura BacterianaKaty 4684Pas encore d'évaluation
- LINFOCITOS T DiapositivasDocument28 pagesLINFOCITOS T DiapositivasBrayan EscobarPas encore d'évaluation
- Patologia Inflamacion Mapa Conceptual. SaavedraDocument11 pagesPatologia Inflamacion Mapa Conceptual. SaavedraJazmin Saavedra Mendoza86% (14)
- InflamaciónDocument9 pagesInflamaciónJosé Gabriel Quiroga Villagra100% (1)
- Tipos de NecrosisDocument33 pagesTipos de Necrosismariesther75% (4)
- Fisiologia Regulaciondelarespiracion 140916092221 Phpapp01Document2 pagesFisiologia Regulaciondelarespiracion 140916092221 Phpapp01Fabiola SanchezPas encore d'évaluation
- Rifampicina Power PointDocument6 pagesRifampicina Power PointKe GuillePas encore d'évaluation
- Sistema Inmune Glosario PDFDocument7 pagesSistema Inmune Glosario PDFCAMILA VARGASPas encore d'évaluation
- ASPERGILOSISDocument7 pagesASPERGILOSISNalgis MendinuetaPas encore d'évaluation
- Inmunidad frente a parásitosDocument16 pagesInmunidad frente a parásitosyccardenasm100% (1)
- Linfocitos BDocument38 pagesLinfocitos Blady laura restrepoPas encore d'évaluation
- Inmunologia t7 Presentacion Antigenica PDFDocument4 pagesInmunologia t7 Presentacion Antigenica PDFYosky AlcPas encore d'évaluation
- Caso VPHDocument23 pagesCaso VPHjeancarlosbazaPas encore d'évaluation
- Sistema Del Complemento: Vías de ActivaciónDocument6 pagesSistema Del Complemento: Vías de ActivaciónDaniela PoncePas encore d'évaluation
- Adaptacion Celular - Robbins!Document14 pagesAdaptacion Celular - Robbins!StephcitaPas encore d'évaluation
- Sistema Del ComplementoDocument19 pagesSistema Del ComplementoSergio David Salazar Lemus100% (1)
- Autoinmunidad. Concepto. Factores Etiopatogénicos. Mecanismos. ClasificaciónDocument5 pagesAutoinmunidad. Concepto. Factores Etiopatogénicos. Mecanismos. ClasificaciónrenzonoePas encore d'évaluation
- PREGUNTAS DE CONTROL GLUCOLISIS - Maria LuizaDocument2 pagesPREGUNTAS DE CONTROL GLUCOLISIS - Maria Luizacharles aires100% (1)
- Cap1 Farmacologia de KatzungDocument8 pagesCap1 Farmacologia de Katzunglucia hunPas encore d'évaluation
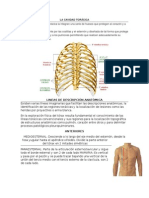
- Lineas de Referencia de TóraxDocument3 pagesLineas de Referencia de TóraxEduardo Navarrete100% (1)
- Inmunodeficiencia Común VariableDocument16 pagesInmunodeficiencia Común VariableJuan HernandezPas encore d'évaluation
- Capitulo 1 de Robbins Respuestas Celulares Ante El Estres y Agreciones Por Toxico Adapataciones, Lesion y MuerteDocument8 pagesCapitulo 1 de Robbins Respuestas Celulares Ante El Estres y Agreciones Por Toxico Adapataciones, Lesion y MuerteFernanda Pineda Gea100% (1)
- Mecanismos Efectores de La Respuesta Inmune InnataDocument7 pagesMecanismos Efectores de La Respuesta Inmune InnataEmilis MadridPas encore d'évaluation
- Señalización Celular y Transducción de Señales FinalDocument7 pagesSeñalización Celular y Transducción de Señales FinalLeón LopezPas encore d'évaluation
- Diéresis y disección de tejidosDocument25 pagesDiéresis y disección de tejidosBETTY FLORES CASTILLOPas encore d'évaluation
- Perfil EnaDocument15 pagesPerfil EnaXavier Emerson Bayona Figueroa50% (4)
- Resumen de LeucemiasDocument9 pagesResumen de LeucemiasDulce SalazarPas encore d'évaluation
- Factores de Coagulacion PDFDocument8 pagesFactores de Coagulacion PDFerikamb54Pas encore d'évaluation
- Mediadores de la inflamación aguda y crónicaDocument20 pagesMediadores de la inflamación aguda y crónicaCristian Jumbo100% (1)
- Glosario de InmunologíaDocument6 pagesGlosario de InmunologíaAngélica SChiPas encore d'évaluation
- Cap 81 y 82 Guyton y Hall: funciones reproductoras masculinas y femeninasDocument9 pagesCap 81 y 82 Guyton y Hall: funciones reproductoras masculinas y femeninasAlexander Gallardo100% (1)
- Capítulo 6 Sistema Del ComplementoDocument26 pagesCapítulo 6 Sistema Del ComplementoJonatan Pinto100% (1)
- Sistema de ComplementoDocument9 pagesSistema de ComplementoLauraCarolinaRestrepoPas encore d'évaluation
- Desnaturalization of The Drugs Expected in The Angela Daniela Drug Yacuanquer Period 2016 - 2017Document4 pagesDesnaturalization of The Drugs Expected in The Angela Daniela Drug Yacuanquer Period 2016 - 2017Conde GustavPas encore d'évaluation
- Cuestionario-14 VIRUSDocument6 pagesCuestionario-14 VIRUSJheanPaulPas encore d'évaluation
- La Muerte y El Duelo en La HipermodernidadDocument6 pagesLa Muerte y El Duelo en La Hipermodernidadhistoriadelperrodinamita100% (1)
- Factores de Riesgo Conductas/Estilo de Vida: Tener Relaciones Sexuales Sin Protección, Ser Sexualmente Activo A Temprana EdadDocument2 pagesFactores de Riesgo Conductas/Estilo de Vida: Tener Relaciones Sexuales Sin Protección, Ser Sexualmente Activo A Temprana EdadCamila Graciela TENORIO TITOPas encore d'évaluation
- Manual Cuidados Auxiliares Básicos PDFDocument352 pagesManual Cuidados Auxiliares Básicos PDFanzacs4742100% (1)
- HistoriaClínicaDocument24 pagesHistoriaClínicaDayana GonzálezPas encore d'évaluation
- Caso Enfoque CognitivoDocument2 pagesCaso Enfoque CognitivoANA MATILDE BIEBERACH VANEGAS100% (1)
- Enfermedades ContagiosasDocument25 pagesEnfermedades Contagiosaslucy DavanPas encore d'évaluation
- Fumarato FerrosoDocument3 pagesFumarato Ferrosoanmich100% (3)
- Trastornos mentales orgánicos: causas y clasificaciónDocument25 pagesTrastornos mentales orgánicos: causas y clasificaciónDULCE ANABELLA DIONICIO LANPas encore d'évaluation
- Guia-Calculo-Cotización DS67Document8 pagesGuia-Calculo-Cotización DS67samuel aguilar riosPas encore d'évaluation
- INFECTOLOGIADocument335 pagesINFECTOLOGIAJorge Nuñez YongPas encore d'évaluation
- Farmacologia Del Sistema Digestivo1Document22 pagesFarmacologia Del Sistema Digestivo1Alexandra MuozPas encore d'évaluation
- Examen obstétrico puérperaDocument13 pagesExamen obstétrico puérperanayareth tapiaPas encore d'évaluation
- Entrevista NeuropsicológicaDocument2 pagesEntrevista Neuropsicológicaramiro50% (2)
- AnticonceptivosDocument5 pagesAnticonceptivosFlorencia RusnokPas encore d'évaluation
- Harrison Manual de Medicina GASTRITIS EROSIVADocument1 pageHarrison Manual de Medicina GASTRITIS EROSIVAAnnery PaulinoPas encore d'évaluation
- Cefalea PostpuncionDocument12 pagesCefalea PostpuncionYaree MorgadoPas encore d'évaluation
- Intoxicaciones AgudasDocument33 pagesIntoxicaciones Agudaswww.pacourgencias.blogspot.com/Pas encore d'évaluation
- NOM 005 Planificación FamiliarDocument38 pagesNOM 005 Planificación FamiliarIto-kun VasquezPas encore d'évaluation
- Asepsia y antisepsia en quirófanoDocument18 pagesAsepsia y antisepsia en quirófanoLucero LoverPas encore d'évaluation
- Gestión de CamasDocument13 pagesGestión de Camasadriana111097Pas encore d'évaluation
- Guía obstétrica V semestreDocument45 pagesGuía obstétrica V semestreKimberly Vanessa HanccoqPas encore d'évaluation
- Enfermedades de la vista y el olfato: causas, síntomas y tratamientosDocument20 pagesEnfermedades de la vista y el olfato: causas, síntomas y tratamientosGénesis Rondón100% (1)
- 12 Manual de Admision FinalDocument142 pages12 Manual de Admision FinalBerrio Arrieta Ovidio JosePas encore d'évaluation
- Farmacos TuberculosisDocument1 pageFarmacos TuberculosisValeria Ticona ChambiPas encore d'évaluation
- Trastorno Infanto-Juveniles Tarea 4Document6 pagesTrastorno Infanto-Juveniles Tarea 4Joseiry Bruno RosarioPas encore d'évaluation
- Homeopatia y Esencias FloralesDocument45 pagesHomeopatia y Esencias FloralesrotzilaPas encore d'évaluation
- Infarto agudo de miocardio: tratamiento y pronósticoDocument11 pagesInfarto agudo de miocardio: tratamiento y pronósticogonzycomPas encore d'évaluation
- 2019 06 26 PGR TH 006 v2 Programa Riesgo PsicosocialDocument26 pages2019 06 26 PGR TH 006 v2 Programa Riesgo PsicosocialJosé MartínezPas encore d'évaluation