Vous aimerez peut-être aussi
- Lipasa Termotolerante de Penicillium SPDocument14 pagesLipasa Termotolerante de Penicillium SPAnonymous XsZi9A5Pas encore d'évaluation
- Sistemas de Apoyo Critico GASESDocument19 pagesSistemas de Apoyo Critico GASESSergio Oswaldo Paniagua MontoyaPas encore d'évaluation
- Método de Kjeldahl en LecheDocument3 pagesMétodo de Kjeldahl en LecheLaura Cristina MoralesPas encore d'évaluation
- Requisitos y clasificación de excipientes farmacéuticosDocument10 pagesRequisitos y clasificación de excipientes farmacéuticosMilagros Santamaría InoñanPas encore d'évaluation
- Actividad Practica 3Document6 pagesActividad Practica 3ISABEL CINTHIA BUSTOS PEÑAPas encore d'évaluation
- Práctica 4Document5 pagesPráctica 4Diego Padilla Santos100% (1)
- Determinación de cenizas en alimentosDocument12 pagesDeterminación de cenizas en alimentosYOBANA ROJASPas encore d'évaluation
- Alcaloides (1) .. FINALDocument11 pagesAlcaloides (1) .. FINALcarlos19895Pas encore d'évaluation
- Practica 13 - MezclasDocument3 pagesPractica 13 - Mezclassandra maria oroya sanchez100% (2)
- Loratadina, el antihistamínico de segunda generaciónDocument9 pagesLoratadina, el antihistamínico de segunda generaciónStefany CáceresPas encore d'évaluation
- Practica 12Document3 pagesPractica 12José A. CóndorPas encore d'évaluation
- Ketoconazol AnalisisDocument2 pagesKetoconazol Analisischiw.Pas encore d'évaluation
- Trabajo de FarmacologiaDocument17 pagesTrabajo de Farmacologiapiero0% (1)
- Practica 4 Saponificacion de Una Grasa PDFDocument7 pagesPractica 4 Saponificacion de Una Grasa PDFCielo LoayzaPas encore d'évaluation
- Analisis Cualitativo y Cuantitativo de La OrfenadrinaDocument7 pagesAnalisis Cualitativo y Cuantitativo de La OrfenadrinaMarco Antonio Tarazona MoyaPas encore d'évaluation
- Farmacoquímica examen finalDocument4 pagesFarmacoquímica examen finalWindows 7Pas encore d'évaluation
- Práctica N 9Document20 pagesPráctica N 9Kalin Kast GonzPas encore d'évaluation
- CLASE 12 PRUEBAS DE FUNCION HEPATICA Practica 2Document17 pagesCLASE 12 PRUEBAS DE FUNCION HEPATICA Practica 2Mari QcPas encore d'évaluation
- Analisis Clinico Tema 2Document38 pagesAnalisis Clinico Tema 2johnPas encore d'évaluation
- Hespiridina NaranjaDocument5 pagesHespiridina NaranjaGerifaltePas encore d'évaluation
- Análisis cuantitativo de AINEsDocument43 pagesAnálisis cuantitativo de AINEsBC NandoPas encore d'évaluation
- 5 - Glucogenolisis y Glucogenogenesis BM e IADocument40 pages5 - Glucogenolisis y Glucogenogenesis BM e IAPaloma SusanPas encore d'évaluation
- Informe 2Document12 pagesInforme 2janssenPas encore d'évaluation
- Marco Teorico Del FluconazolDocument1 pageMarco Teorico Del Fluconazolyassury jannet lluen llontopPas encore d'évaluation
- Practica N°9 y 10Document4 pagesPractica N°9 y 10VEN2 RAPas encore d'évaluation
- Ensayo de Gineco Diabetes GestacionalDocument12 pagesEnsayo de Gineco Diabetes GestacionalDaniela D. Ramirez Magaña0% (1)
- Absorción de Drogas Desde El Tracto GastrointestinalDocument12 pagesAbsorción de Drogas Desde El Tracto GastrointestinalJUAN PABLO CORONA MONTUFARPas encore d'évaluation
- Informe de Cefradina y Furosemida 1Document18 pagesInforme de Cefradina y Furosemida 1Stefany CáceresPas encore d'évaluation
- Análisis cualitativo etoricoxibDocument12 pagesAnálisis cualitativo etoricoxibEstefany Quispe Ramirez100% (1)
- Evolución histórica de la química farmacéuticaDocument5 pagesEvolución histórica de la química farmacéuticaPedro CampoPas encore d'évaluation
- Formato de Practica de ClorfenaminaDocument3 pagesFormato de Practica de Clorfenaminajanet huanca choquePas encore d'évaluation
- Cuestionario Bioquimica PracticaDocument3 pagesCuestionario Bioquimica PracticaNayely Arrieta GarcíaPas encore d'évaluation
- Sistema de Clasificación BiofarmaceúticaDocument2 pagesSistema de Clasificación Biofarmaceúticaastrid7_berenice7Pas encore d'évaluation
- Informe CefalosporinaDocument13 pagesInforme CefalosporinaEduardo MalpartidaPas encore d'évaluation
- Purinas y Pirimidinas 2010Document81 pagesPurinas y Pirimidinas 2010joe_fox2132296Pas encore d'évaluation
- Cuestionario AtenololDocument2 pagesCuestionario AtenololDiego Carrion AlvaPas encore d'évaluation
- Examen Parcial - FCDocument11 pagesExamen Parcial - FC12-4560% (1)
- Protocolo Productos CarnicosDocument4 pagesProtocolo Productos Carnicosfabian jaraPas encore d'évaluation
- Farmacomorfica Lab.3Document7 pagesFarmacomorfica Lab.3javier0% (1)
- Análisis fitoquímico de extractos de Eucalipto, Romero y BorracheroDocument12 pagesAnálisis fitoquímico de extractos de Eucalipto, Romero y BorracheroPaolaMoraPas encore d'évaluation
- Factores que influyen en la absorción de fármacosDocument17 pagesFactores que influyen en la absorción de fármacosDiego APas encore d'évaluation
- Sistemas de Apoyo CriticoDocument5 pagesSistemas de Apoyo CriticoVictoria Perez100% (1)
- Introduccion - Marco TeoricoDocument7 pagesIntroduccion - Marco TeoricohillaryPas encore d'évaluation
- Procedimiento PreparacionDocument2 pagesProcedimiento PreparacionMaria Joa Luyo AntonioPas encore d'évaluation
- Métodos de secado farmacéuticoDocument11 pagesMétodos de secado farmacéuticoYeseniaPas encore d'évaluation
- LoratadinaDocument10 pagesLoratadinaStefany CáceresPas encore d'évaluation
- Hematimetría: Recuento de glóbulos rojos, hemoglobina y hematocritoDocument2 pagesHematimetría: Recuento de glóbulos rojos, hemoglobina y hematocritoLiliana Miranda APas encore d'évaluation
- Bromatologia Informe 2Document7 pagesBromatologia Informe 2williamPas encore d'évaluation
- Choke 2017Document48 pagesChoke 2017Joseph AngelPas encore d'évaluation
- HCTZ 25 MG Una-WPS OfficeDocument2 pagesHCTZ 25 MG Una-WPS OfficeSantiagoDazaPas encore d'évaluation
- PRM5 DepresionDocument15 pagesPRM5 DepresionMarco Mamani HuamanculiPas encore d'évaluation
- CuestionarioDocument4 pagesCuestionariomaikolPas encore d'évaluation
- FB7M3 - Farmacologia 2Document3 pagesFB7M3 - Farmacologia 2Miguel Milano FloresPas encore d'évaluation
- INFORME QF II CiprofloxacinoDocument7 pagesINFORME QF II Ciprofloxacinocristian david hurtado peñaPas encore d'évaluation
- Pasito Tecnologia 3 2022 I - Francesca RojasDocument4 pagesPasito Tecnologia 3 2022 I - Francesca RojasFrancesca Rojas100% (1)
- Practica n2 FarmacoquimjcaDocument3 pagesPractica n2 FarmacoquimjcaAlvaro Rolando Paredes TacoPas encore d'évaluation
- IMA-medicmentos Que Afectan La IngestaDocument38 pagesIMA-medicmentos Que Afectan La IngestaSaphiraLainPas encore d'évaluation
- 02 Mothe and Reddy 2016 - Production Purification and Characterization-Protease-Bacillus - En.esDocument10 pages02 Mothe and Reddy 2016 - Production Purification and Characterization-Protease-Bacillus - En.esArturo PeñablassiPas encore d'évaluation
- BioingenieríaDocument8 pagesBioingenieríajesusPas encore d'évaluation
- Enzimas As Por HongosDocument6 pagesEnzimas As Por HongosFelix BustamantePas encore d'évaluation
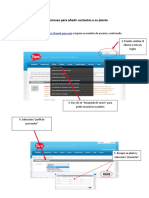
- Instrucciones para Añadir Contactos A Su Planta (SPN)Document3 pagesInstrucciones para Añadir Contactos A Su Planta (SPN)Alexander UFPas encore d'évaluation
- Costeo por órdenes de producciónDocument10 pagesCosteo por órdenes de producciónAlexander UFPas encore d'évaluation
- Normas de Presentación de Los Trabajos de Investigación Y Tesis La Molina, Lima Perú Abril 2019Document37 pagesNormas de Presentación de Los Trabajos de Investigación Y Tesis La Molina, Lima Perú Abril 2019Alexander UFPas encore d'évaluation
- Ficha Técnica CAJUDocument2 pagesFicha Técnica CAJUAlexander UFPas encore d'évaluation
- Guia - Costos y Presupuestos I PDFDocument44 pagesGuia - Costos y Presupuestos I PDFAlexander UFPas encore d'évaluation
- Dialnet OptimizacionDeLaConcentracionDeLaAmilasaYLactosuer 6583429Document10 pagesDialnet OptimizacionDeLaConcentracionDeLaAmilasaYLactosuer 6583429Sugar DaddyPas encore d'évaluation
- Análisis CVU punto equilibrioDocument36 pagesAnálisis CVU punto equilibrio2212leoPas encore d'évaluation
- Tesis en Ciencias Contables PDFDocument112 pagesTesis en Ciencias Contables PDFcosmelPas encore d'évaluation
- Menestras Del Peru para El MundoDocument92 pagesMenestras Del Peru para El MundoAlexander UF100% (2)
- Sistema de Inocuidad en Los AlimentosDocument62 pagesSistema de Inocuidad en Los AlimentosDavid PalmarPas encore d'évaluation
- Plantilla Diccionario - WBSDocument2 pagesPlantilla Diccionario - WBSAlexander UFPas encore d'évaluation
- BPM - 21 CFR 110Document15 pagesBPM - 21 CFR 110Alexander UFPas encore d'évaluation
- Referencias para El Nivel de Cloro Libre Residual en El AguaDocument1 pageReferencias para El Nivel de Cloro Libre Residual en El AguaAlexander UFPas encore d'évaluation
- 2.1+AIB+Consolidated+Standards+Food Contact+Packaging EspDocument51 pages2.1+AIB+Consolidated+Standards+Food Contact+Packaging EspmariajsalasPas encore d'évaluation
- Referencias para El Nivel de Cloro Libre Residual en El AguaDocument1 pageReferencias para El Nivel de Cloro Libre Residual en El AguaAlexander UFPas encore d'évaluation
- Referencias para El Nivel de Cloro Libre Residual en El AguaDocument1 pageReferencias para El Nivel de Cloro Libre Residual en El AguaAlexander UFPas encore d'évaluation
- Aditivos Alimentarios-Laboratorios-Normativa Enac-Microbiolo PDFDocument252 pagesAditivos Alimentarios-Laboratorios-Normativa Enac-Microbiolo PDFAlexander UFPas encore d'évaluation
- Analisis de La UbicaciónDocument1 pageAnalisis de La UbicaciónAlexander UFPas encore d'évaluation
- D.A. 093 Modif To Ordenanza 295 - 28 - Feb - 2003Document5 pagesD.A. 093 Modif To Ordenanza 295 - 28 - Feb - 2003Richard FigueroaPas encore d'évaluation
- Informe 1Document110 pagesInforme 1Alexander UFPas encore d'évaluation
- Sasonadores y EspaciesDocument195 pagesSasonadores y EspaciesAlexander UFPas encore d'évaluation
- 122 Tes Is Alejandra S IqDocument113 pages122 Tes Is Alejandra S IqAlexander UFPas encore d'évaluation
- 7 Nuevas herramientas para mejorar calidadDocument13 pages7 Nuevas herramientas para mejorar calidadAlexander UFPas encore d'évaluation
- Industria Del MalteadoDocument4 pagesIndustria Del MalteadoAlexander UFPas encore d'évaluation
- Mango ProximidadDocument2 pagesMango ProximidadAlexander UFPas encore d'évaluation
- GCen La Univer de RiojaDocument7 pagesGCen La Univer de RiojaAlexander UFPas encore d'évaluation
- Agitacion 2013Document21 pagesAgitacion 2013Alexander UFPas encore d'évaluation
- Informe de Pruebas en PanDocument6 pagesInforme de Pruebas en PanAlexander UFPas encore d'évaluation
- GCen La Univer de RiojaDocument7 pagesGCen La Univer de RiojaAlexander UFPas encore d'évaluation
- COMPRESIONDocument24 pagesCOMPRESIONAlexander UFPas encore d'évaluation
- Tema 5 Enzimas y VitaminasDocument13 pagesTema 5 Enzimas y VitaminasDavid MorenoPas encore d'évaluation
- Beta Oxidación 1Document11 pagesBeta Oxidación 1Anonymous TaDw7ejHk100% (1)
- Bases Biologicas de La Vida CVCTGDocument8 pagesBases Biologicas de La Vida CVCTGMagali González100% (3)
- Distribución de cromosomas durante la división celularDocument20 pagesDistribución de cromosomas durante la división celularSofía UrbinaPas encore d'évaluation
- Pared CelularDocument5 pagesPared CelularColumba LopezPas encore d'évaluation
- Biomoleculas IntroduccionDocument5 pagesBiomoleculas IntroduccionSantiago SalazarPas encore d'évaluation
- Bioenergética 2 2012 UNABDocument57 pagesBioenergética 2 2012 UNABPaTricia GonzalezPas encore d'évaluation
- Conceptos Básicos Sobre InmunologíaDocument25 pagesConceptos Básicos Sobre InmunologíaJc Loz100% (3)
- Solucionario: Glicerina + 3 Ác. Palmítico Tripalmitina + 3 AguaDocument7 pagesSolucionario: Glicerina + 3 Ác. Palmítico Tripalmitina + 3 AguaGustavo SergioPas encore d'évaluation
- Preguntas de Repaso, Arumir Ortiz Adriana MayteDocument7 pagesPreguntas de Repaso, Arumir Ortiz Adriana MayteArumir OrtizPas encore d'évaluation
- Ciclo celular y sus fases claveDocument35 pagesCiclo celular y sus fases claveWalter GUILLEN POZOPas encore d'évaluation
- Identificación de Enlaces PeptidocosDocument3 pagesIdentificación de Enlaces Peptidocosimar555Pas encore d'évaluation
- TEMA 4 ProteinasDocument13 pagesTEMA 4 ProteinasDanielys PerezPas encore d'évaluation
- Introducción A Los Procesos de BioseparaciónDocument70 pagesIntroducción A Los Procesos de BioseparaciónJuan Jesús Cruz MaldonadoPas encore d'évaluation
- Farmacocinética AbsorciónDocument4 pagesFarmacocinética AbsorciónJavier VMPas encore d'évaluation
- Farmacologia - Receptores FarmacológicosDocument65 pagesFarmacologia - Receptores FarmacológicosMarce811882% (11)
- Resumen Del NucleoDocument2 pagesResumen Del NucleoSilvia Leiva FierroPas encore d'évaluation
- Fisiopatología y Opciones de Tratamiento Futuro La Encefalopatía HepáticaDocument9 pagesFisiopatología y Opciones de Tratamiento Futuro La Encefalopatía HepáticaSerendipiaPas encore d'évaluation
- 2021Document76 pages2021Yousef A.Pas encore d'évaluation
- Farmacologia SimpaticomimeticosDocument16 pagesFarmacologia SimpaticomimeticosGuadalupe Jojo100% (1)
- Segundo MapaDocument1 pageSegundo MapaEstrella López TorresPas encore d'évaluation
- Wuolah Free BIOLOGIA MOLECULAR TEMA 1Document23 pagesWuolah Free BIOLOGIA MOLECULAR TEMA 1Maria111200Pas encore d'évaluation
- Guia de Laboratorio EnzimasDocument3 pagesGuia de Laboratorio EnzimasNalda Pillco ChicllaPas encore d'évaluation
- INJURIA CELULAR Dra JaraDocument57 pagesINJURIA CELULAR Dra JaraEdward ChacmaPas encore d'évaluation
- Agar LIADocument2 pagesAgar LIArichiiehzu100% (1)
- Paredes celulares hongos, algas, arqueas y bacteriasDocument3 pagesParedes celulares hongos, algas, arqueas y bacteriasBrandonCaballeroPas encore d'évaluation
- Trastornos metabolismo carbohidratosDocument20 pagesTrastornos metabolismo carbohidratosJuan CastroPas encore d'évaluation
- Semana 2-Guion de Video-Los NeurotransmisoresDocument4 pagesSemana 2-Guion de Video-Los NeurotransmisoresNicole EstradaPas encore d'évaluation
- EsternoideDocument2 pagesEsternoideanibalPas encore d'évaluation
- GLUCIDOSDocument18 pagesGLUCIDOSleoncio lenciPas encore d'évaluation