Vous aimerez peut-être aussi
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (121)
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (588)
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (400)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (266)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5794)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2259)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (345)
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (895)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (74)
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- Electro-Etching Made EasyDocument52 pagesElectro-Etching Made Easyedhykoes100% (1)
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- Phytochemicals: Extraction Methods, Basic Structures and Mode of Action As Potential Chemotherapeutic AgentsDocument33 pagesPhytochemicals: Extraction Methods, Basic Structures and Mode of Action As Potential Chemotherapeutic AgentsMacDonald OkwudiliChukwu Stain100% (1)
- Steam BookDocument66 pagesSteam Bookdekate_manoj100% (1)
- DS4B - (1988) Physical Constants of Hydrocarbon and Non-Hydrocarbon CompoundsDocument216 pagesDS4B - (1988) Physical Constants of Hydrocarbon and Non-Hydrocarbon CompoundsJacques Strappe0% (1)
- Induction FurnaceDocument16 pagesInduction FurnaceMURSELIM ALIPas encore d'évaluation
- Physical Progress Weightage Rev 03Document1 pagePhysical Progress Weightage Rev 03burereyPas encore d'évaluation
- What Is ChemistryDocument26 pagesWhat Is ChemistryAnonymousGodiswithyouPas encore d'évaluation
- Supplement 10 TS Calibration ECSPP ECBSDocument16 pagesSupplement 10 TS Calibration ECSPP ECBSKingston Rivington100% (1)
- Previous View Home: Dimensions Performance SpecificationDocument124 pagesPrevious View Home: Dimensions Performance SpecificationIntan HaddadPas encore d'évaluation
- Article 7 Teacher As Actor: Future English Teachers' Training in The Natural Approach Using DramaDocument13 pagesArticle 7 Teacher As Actor: Future English Teachers' Training in The Natural Approach Using DramaIntan HaddadPas encore d'évaluation
- Lateral Lingual Swellings and One Medial Swelling, The Tuberculum ImparDocument1 pageLateral Lingual Swellings and One Medial Swelling, The Tuberculum ImparIntan HaddadPas encore d'évaluation
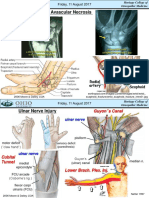
- Scaphoid Fracture & Avascular Necrosis: Friday, 11 August 2017Document4 pagesScaphoid Fracture & Avascular Necrosis: Friday, 11 August 2017Intan HaddadPas encore d'évaluation
- Effect of Physiotherapy Rehabilitation On Volkmann Ischemic Contracture-A Case StudyDocument2 pagesEffect of Physiotherapy Rehabilitation On Volkmann Ischemic Contracture-A Case StudyIntan HaddadPas encore d'évaluation
- 2003 Dot Itp EhtfDocument11 pages2003 Dot Itp EhtfNoirchevalPas encore d'évaluation
- CHEM 1067 Lec 3 - 2019 - NJ - 4 PDFDocument21 pagesCHEM 1067 Lec 3 - 2019 - NJ - 4 PDFIbrahim AliPas encore d'évaluation
- One-Dimensional Compressible Flow in Variable AreaDocument6 pagesOne-Dimensional Compressible Flow in Variable AreaswatiPas encore d'évaluation
- Class 12 Chemistry Syllabus 2023-24Document3 pagesClass 12 Chemistry Syllabus 2023-24Rooh KSHIVPas encore d'évaluation
- Synthesis of Paracetamol by Liquid Phase Beckmann Rearrangement of 4-Hydroxyacetophenone Oxime Over H3PO4/Al-MCM-41Document8 pagesSynthesis of Paracetamol by Liquid Phase Beckmann Rearrangement of 4-Hydroxyacetophenone Oxime Over H3PO4/Al-MCM-41MUHAMMAD RIDHO ANSYARIPas encore d'évaluation
- Ejercicios AliviosDocument20 pagesEjercicios AliviosPaola ChavezPas encore d'évaluation
- Industrial Types of Gold Deposits of The East KazakhstanDocument9 pagesIndustrial Types of Gold Deposits of The East KazakhstanGEOLINKS International Conference 2019Pas encore d'évaluation
- Fine Bubble aeration-EPADocument7 pagesFine Bubble aeration-EPARizkydwiArizkaPas encore d'évaluation
- Lab Waste ManagementDocument29 pagesLab Waste ManagementMohamed AslamPas encore d'évaluation
- Chemical Digestion and Absorption - A Closer Look - Anatomy and PhysiologyDocument20 pagesChemical Digestion and Absorption - A Closer Look - Anatomy and PhysiologyMa Mayla Imelda LapaPas encore d'évaluation
- Study of Cement Treated Base Agreagate Properties For Pavement StructureDocument9 pagesStudy of Cement Treated Base Agreagate Properties For Pavement StructureOrlando RojasPas encore d'évaluation
- Material Safety AvabexDocument4 pagesMaterial Safety Avabexfs1640Pas encore d'évaluation
- SEJ446 Final Report 2014Document43 pagesSEJ446 Final Report 2014Busiku SilengaPas encore d'évaluation
- Alfa 50 RegulatorDocument2 pagesAlfa 50 Regulatorykoze100% (1)
- 4 Composite Solid PropellantDocument18 pages4 Composite Solid PropellantAli KhanPas encore d'évaluation
- As 1221 Hose-ReelsDocument2 pagesAs 1221 Hose-ReelsDo BuiPas encore d'évaluation
- Drop Test TitrationDocument2 pagesDrop Test TitrationHadid SukmanaPas encore d'évaluation
- End Term Exam Chemistry Year 13Document13 pagesEnd Term Exam Chemistry Year 13Barminga KamurenPas encore d'évaluation
- ST2 PDFDocument5 pagesST2 PDFKaraline MarcesPas encore d'évaluation
- 5lab - Serial DilutionDocument3 pages5lab - Serial DilutionBianca APas encore d'évaluation
- Explanation Text - Reading Comprehensive - Text 1 To 3Document4 pagesExplanation Text - Reading Comprehensive - Text 1 To 3DEA ANNPas encore d'évaluation
- Extraction and Characterization of Hemicellulose From The Corn Fiber Produced by Corn Wet-Milling ProcessesDocument5 pagesExtraction and Characterization of Hemicellulose From The Corn Fiber Produced by Corn Wet-Milling ProcessesmbolantenainaPas encore d'évaluation
- Jotun Thinner No. 2 - EnglishDocument13 pagesJotun Thinner No. 2 - EnglishBams ArifinPas encore d'évaluation