Vous aimerez peut-être aussi
- Fisheries ENTREPLecture From Mam HelenDocument176 pagesFisheries ENTREPLecture From Mam Helengriffith jamesPas encore d'évaluation
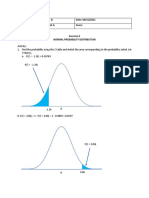
- X 213231124Document23 pagesX 213231124Seave HwangPas encore d'évaluation
- Document 1Document38 pagesDocument 1gosaye desalegnPas encore d'évaluation
- Farm Management Case Study (Revised From Hillier and Lieberman's Introduction To Operations Research)Document3 pagesFarm Management Case Study (Revised From Hillier and Lieberman's Introduction To Operations Research)rssarinPas encore d'évaluation
- Inventory Control Subject To Known DemandDocument37 pagesInventory Control Subject To Known DemandAfifahSeptiaPas encore d'évaluation
- Elasticity Practice TestDocument8 pagesElasticity Practice TestAmir NelsonPas encore d'évaluation
- Leno Company Makes Swimsuits and Sells These Suits Directly ToDocument1 pageLeno Company Makes Swimsuits and Sells These Suits Directly ToMuhammad ShahidPas encore d'évaluation
- A FisherDocument2 pagesA FisherAngel Mae AcuestaPas encore d'évaluation
- 2.5 Consumer and Producer Surplus - Phuong - KeyDocument15 pages2.5 Consumer and Producer Surplus - Phuong - KeyMy Fair LadyPas encore d'évaluation
- Chapter 18-Practice ExsercisesDocument18 pagesChapter 18-Practice ExsercisesThiPas encore d'évaluation
- Quiz 3Document1 pageQuiz 3Marc ViduyaPas encore d'évaluation
- CMA-Unit 3-Costing of Materials 18-19Document48 pagesCMA-Unit 3-Costing of Materials 18-19RahulPas encore d'évaluation
- Chapter # 7 Exercise & ProblemsDocument4 pagesChapter # 7 Exercise & ProblemsZia Uddin0% (1)
- Managerial 1Document88 pagesManagerial 1Mary Kris CaparosoPas encore d'évaluation
- Mang Isko and The Tragedy of The CommonsDocument2 pagesMang Isko and The Tragedy of The CommonsSophia Cianna FaderugaoPas encore d'évaluation
- STS-Midterm ExaminationDocument2 pagesSTS-Midterm ExaminationGeonyzl L. AlviolaPas encore d'évaluation
- Economics: Principles in Action NOTES For Chapter 1Document1 pageEconomics: Principles in Action NOTES For Chapter 1Sebastian RamirezPas encore d'évaluation
- Climate Change Affects BiodiversityDocument8 pagesClimate Change Affects BiodiversityAlexander MccormickPas encore d'évaluation
- The Cost of Goods Manufactured ScheduleDocument7 pagesThe Cost of Goods Manufactured ScheduleM Jamal KhanPas encore d'évaluation
- Exam Questions and AnswersDocument18 pagesExam Questions and AnswersAna Aliyah Angin100% (1)
- Sample Questions - Accounting For OverheadDocument4 pagesSample Questions - Accounting For OverheadRedPas encore d'évaluation
- QUIZ 9 - Chapters 9 and 10: ECONOMIC DEVELOPMENT - Finals (Summer)Document5 pagesQUIZ 9 - Chapters 9 and 10: ECONOMIC DEVELOPMENT - Finals (Summer)sofiaPas encore d'évaluation
- TVMPractice ProbDocument6 pagesTVMPractice ProbShaikh Shahrukh FarhanPas encore d'évaluation
- Ch24 ProblemsDocument33 pagesCh24 ProblemsWed CornelPas encore d'évaluation
- EdX Stats ReadingsDocument244 pagesEdX Stats Readingsseggy7100% (1)
- FIN 205 Tutorial Questions Sem 3 2021Document55 pagesFIN 205 Tutorial Questions Sem 3 2021黄于绮Pas encore d'évaluation
- LP Maximization Graphical Method 2021Document28 pagesLP Maximization Graphical Method 2021liesly buticPas encore d'évaluation
- Assignment 4 E OfficeDocument1 pageAssignment 4 E OfficeJehan BimbasPas encore d'évaluation
- The 2017 PICE VSU SC Constitution and By-LawsDocument14 pagesThe 2017 PICE VSU SC Constitution and By-LawsJohn GaviolaPas encore d'évaluation
- Operations Research Tutorial ProblemDocument4 pagesOperations Research Tutorial ProblemAjitsingh JagtapPas encore d'évaluation
- Ch17 Solution PDFDocument53 pagesCh17 Solution PDFmaria dezarie brioso100% (1)
- BIP-Week-13-14-with NotesDocument8 pagesBIP-Week-13-14-with NotesgelPas encore d'évaluation
- Quiz 4Document6 pagesQuiz 4BibliophilioManiac100% (1)
- F. Reif - Statistical Physics - Chapter 1 PDFDocument51 pagesF. Reif - Statistical Physics - Chapter 1 PDFIndrawati WilujengPas encore d'évaluation
- World Fisheries and Aquaculture: The State ofDocument28 pagesWorld Fisheries and Aquaculture: The State ofAnonymous xv5fUs4AvPas encore d'évaluation
- 6.2gonzales - JM - 2bsma-A - Exercise 6 - NPDDocument4 pages6.2gonzales - JM - 2bsma-A - Exercise 6 - NPDJet jet GonzalesPas encore d'évaluation
- Review of AccountingDocument31 pagesReview of AccountingDylan LowePas encore d'évaluation
- Managerial AccountingMid Term ExaminationDocument3 pagesManagerial AccountingMid Term ExaminationjaePas encore d'évaluation
- MS WS 1 (Breakeven)Document2 pagesMS WS 1 (Breakeven)Omkar Reddy Punuru100% (1)
- Research Proposal Hamza DawidDocument20 pagesResearch Proposal Hamza DawidHamza Dawid HamidPas encore d'évaluation
- GED102 M1 W1 L1 HW A1.2 (Done) PDFDocument2 pagesGED102 M1 W1 L1 HW A1.2 (Done) PDFDaniel RamosPas encore d'évaluation
- Investment in Associate-Mytha Isabel D. SalesDocument9 pagesInvestment in Associate-Mytha Isabel D. SalesMytha Isabel SalesPas encore d'évaluation
- Investment Analysis and Portfolio Management Christmas Worksheet 2009Document2 pagesInvestment Analysis and Portfolio Management Christmas Worksheet 2009farrukhazeemPas encore d'évaluation
- Modeling of Professional and Scientific Staff ManagementDocument13 pagesModeling of Professional and Scientific Staff Managementst57143Pas encore d'évaluation
- P1 QuestionsDocument11 pagesP1 QuestionsjustjadePas encore d'évaluation
- FAP T11 KuralayDocument5 pagesFAP T11 KuralayKuralay TilegenPas encore d'évaluation
- Ch. 9 Production and ProductivityDocument6 pagesCh. 9 Production and ProductivityHANNAH GODBEHEREPas encore d'évaluation
- The Role of Language in GlobalizationDocument3 pagesThe Role of Language in GlobalizationAbhas BhargavaPas encore d'évaluation
- Economics Chapter 4-9Document51 pagesEconomics Chapter 4-9Warwick PangilinanPas encore d'évaluation
- Multiple Choice QuestionsDocument23 pagesMultiple Choice QuestionsKim Audrey Jalalain0% (1)
- 15chap 3.1 Sampling DistributionDocument33 pages15chap 3.1 Sampling DistributionNyah MargarettPas encore d'évaluation
- Module 3 Part 1formulation of LP ProblemsDocument42 pagesModule 3 Part 1formulation of LP Problemshello hahahPas encore d'évaluation
- Fin Acc Exam CompilationDocument27 pagesFin Acc Exam CompilationRiza100% (1)
- (Semi-Variable) Total Variable) Cost As Production Unit Fixed Costs) As Production IncreaseDocument4 pages(Semi-Variable) Total Variable) Cost As Production Unit Fixed Costs) As Production IncreaseClint AbenojaPas encore d'évaluation
- National Income ConceptsDocument15 pagesNational Income Conceptsrinky_trivediPas encore d'évaluation
- Team One Group Final ProjectDocument17 pagesTeam One Group Final Projectdee kee100% (1)
- Triple Benefits From Spatial Resource ManagementDocument8 pagesTriple Benefits From Spatial Resource ManagementRaúl AltamiranoPas encore d'évaluation
- Credit and CottonDocument9 pagesCredit and CottonKhalid MushtaqPas encore d'évaluation
- Consumption And: Unit 7 Asset PricesDocument13 pagesConsumption And: Unit 7 Asset PricesnitikanehiPas encore d'évaluation
- Disturbance Effects On Financial Timberland Returns in AustriaDocument12 pagesDisturbance Effects On Financial Timberland Returns in AustriaShakti DalabeheraPas encore d'évaluation
- Halkola TainaDocument66 pagesHalkola TainaVittorioPas encore d'évaluation
- M4 - E1 - NylPro Case - Solution Presentation PDFDocument14 pagesM4 - E1 - NylPro Case - Solution Presentation PDFVittorioPas encore d'évaluation
- C1 ClarksonBlack - 1Document12 pagesC1 ClarksonBlack - 1VittorioPas encore d'évaluation
- Escp Europe Mim GBDocument8 pagesEscp Europe Mim GBVittorioPas encore d'évaluation
- Lightning Calculators: Dead Reckonings: Lost Art in The Mathematical SciencesDocument31 pagesLightning Calculators: Dead Reckonings: Lost Art in The Mathematical SciencesVittorioPas encore d'évaluation
- MC 2 Consulting BidDocument40 pagesMC 2 Consulting BidbenelovesPas encore d'évaluation
- Labor Relations - Policy and DefinitionsDocument4 pagesLabor Relations - Policy and Definitionschisel_159100% (1)
- IT Governance Mod 5Document40 pagesIT Governance Mod 5Asmita Bhagdikar100% (1)
- Banking&Ins Short Notes-By JurazDocument41 pagesBanking&Ins Short Notes-By JurazIt's me100% (9)
- Target Costing AnsDocument5 pagesTarget Costing Anszoyashaikh20Pas encore d'évaluation
- Solved Nora Transfers To Needle Corporation Depreciable Machinery Originally Costing 18 000Document1 pageSolved Nora Transfers To Needle Corporation Depreciable Machinery Originally Costing 18 000Anbu jaromiaPas encore d'évaluation
- ChapTER 17Document51 pagesChapTER 17Yenelyn Apistar Cambarijan0% (1)
- Module 1 (Law On Sales)Document17 pagesModule 1 (Law On Sales)The Brain Dump PHPas encore d'évaluation
- Equity Fincing RiskDocument62 pagesEquity Fincing RiskBrian HughesPas encore d'évaluation
- Examination: Subject CA1 Core Applications Concepts Paper 1 (Assets)Document3 pagesExamination: Subject CA1 Core Applications Concepts Paper 1 (Assets)Saad MalikPas encore d'évaluation
- Week 8 Entrep (April 26-30)Document7 pagesWeek 8 Entrep (April 26-30)LouissePas encore d'évaluation
- SWOT Analysis of NestleDocument2 pagesSWOT Analysis of NestleMuddasar AbbasiPas encore d'évaluation
- Sol. Man. - Chapter 5 - Corporate Liquidation Reorganization - 2020 EditionDocument22 pagesSol. Man. - Chapter 5 - Corporate Liquidation Reorganization - 2020 Editiondianel villarico100% (1)
- Case 1 - Promoting A Breakfast CerealDocument4 pagesCase 1 - Promoting A Breakfast CerealArpit Chaudhary0% (1)
- 2015 BCom Hons 2015 SemSysSyllabus 030615Document38 pages2015 BCom Hons 2015 SemSysSyllabus 030615kalpesh deora100% (2)
- OJT Report SampleDocument49 pagesOJT Report SampleKimberly Ann Lumanog AmarPas encore d'évaluation
- Butler Lumber Company PDFDocument13 pagesButler Lumber Company PDFFreshela AtasPas encore d'évaluation
- Dividend Policy Theory PDFDocument14 pagesDividend Policy Theory PDFXael AlvarezPas encore d'évaluation
- Document1nsd A JnbasdhbkjhdhbkgehkgfDocument4 pagesDocument1nsd A JnbasdhbkjhdhbkgehkgfAhmed NoorPas encore d'évaluation
- Corporatisation and Demutualisation of Stock Exchanges: I. What Is Corporatis-Ation and Demutuali - Sation?Document5 pagesCorporatisation and Demutualisation of Stock Exchanges: I. What Is Corporatis-Ation and Demutuali - Sation?Ashish UpadhyayaPas encore d'évaluation
- Production and Productivity: Lecture Session 01Document23 pagesProduction and Productivity: Lecture Session 01Charitha RanwalaPas encore d'évaluation
- Solution Manual For College Accounting 22nd EditionDocument18 pagesSolution Manual For College Accounting 22nd EditionMichaelRamseydgjk100% (38)
- The Aim of Marketing Is To Make Selling Superfluous by Understanding The Customer So Well The Product or Service Fits HimDocument8 pagesThe Aim of Marketing Is To Make Selling Superfluous by Understanding The Customer So Well The Product or Service Fits HimAmit MaityPas encore d'évaluation
- 2022 StatementsDocument43 pages2022 StatementssachinPas encore d'évaluation
- Five-Forces Analysis For CSD Cola Wars CDocument2 pagesFive-Forces Analysis For CSD Cola Wars CashePas encore d'évaluation
- Corporate Governance of Aditya Birla NuvoDocument10 pagesCorporate Governance of Aditya Birla NuvoZoheb BagwanPas encore d'évaluation
- Marketing MythsDocument2 pagesMarketing MythsMuhammad Hasbi BaharuddinPas encore d'évaluation
- Module Aa2 Lab (Odd 2021-2022) FinalDocument57 pagesModule Aa2 Lab (Odd 2021-2022) FinalDenisse Aretha LeePas encore d'évaluation
- Cash Flow AnalysisDocument7 pagesCash Flow AnalysisDr. Shoaib MohammedPas encore d'évaluation
- 2-Notes On Banking Products & Services-Part 1Document16 pages2-Notes On Banking Products & Services-Part 1Kirti GiyamalaniPas encore d'évaluation