Vous aimerez peut-être aussi
- Na BenzoDocument30 pagesNa BenzoSiti AsnaniPas encore d'évaluation
- 10 1111@j 1574-695X 1996 tb00251 XDocument5 pages10 1111@j 1574-695X 1996 tb00251 XDuyet TranPas encore d'évaluation
- The Hearing-Loss GuideDocument284 pagesThe Hearing-Loss GuideDuyet Tran100% (1)
- The Hearing-Loss GuideDocument284 pagesThe Hearing-Loss GuideDuyet Tran100% (1)
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5783)
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (587)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (890)
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (72)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (399)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (265)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (344)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2219)
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (119)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- The Beginning of Dr. Jose RizalDocument29 pagesThe Beginning of Dr. Jose RizalEd PascasioPas encore d'évaluation
- Field Study Learning Episode: On Scoring RubricDocument4 pagesField Study Learning Episode: On Scoring Rubricaya camanggocPas encore d'évaluation
- VEE Approved CoursesDocument5 pagesVEE Approved CoursesLetsogile BaloiPas encore d'évaluation
- Lesson 1 Inclusive Education in Special EducationDocument7 pagesLesson 1 Inclusive Education in Special Educationsinco lovelyn rosePas encore d'évaluation
- CAPE Communication Studies ReflectionDocument3 pagesCAPE Communication Studies ReflectionRuel-John CrawfordPas encore d'évaluation
- Jasiora: Kebijakan Pengoperasian Terminal Kota Lintas Muara Bungo: Dinamika Dan PermasalahannyaDocument13 pagesJasiora: Kebijakan Pengoperasian Terminal Kota Lintas Muara Bungo: Dinamika Dan PermasalahannyaIrfan Nurfauzan IskandarPas encore d'évaluation
- MSPE TemplateDocument2 pagesMSPE TemplateAndalibShaikh0% (1)
- Banawa v. Mirano: Is an illegitimate child or adopted child entitled to inherit from relatives of deceasedDocument2 pagesBanawa v. Mirano: Is an illegitimate child or adopted child entitled to inherit from relatives of deceasedJeng PionPas encore d'évaluation
- EDU 32 PreliminariesDocument9 pagesEDU 32 PreliminariesRichee LunnayPas encore d'évaluation
- IB TOK Essay ExampleDocument4 pagesIB TOK Essay ExampleLucy HanPas encore d'évaluation
- Week 2 PHILIPPINE POLITICS AND GOVERNANCE SEPT 20-24-2021Document4 pagesWeek 2 PHILIPPINE POLITICS AND GOVERNANCE SEPT 20-24-2021Taj NgilayPas encore d'évaluation
- College of Arts and Sciences Education GE1 - Course SyllabusDocument8 pagesCollege of Arts and Sciences Education GE1 - Course SyllabusMizzy BaldoPas encore d'évaluation
- Resume of Laminor27Document2 pagesResume of Laminor27api-27481876Pas encore d'évaluation
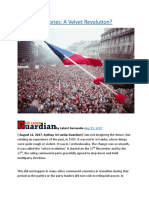
- Occasional Stories A Velvet RevolutionDocument7 pagesOccasional Stories A Velvet RevolutionThavamPas encore d'évaluation
- Ac9 Civ Cit Subject Overview Yr7-10Document20 pagesAc9 Civ Cit Subject Overview Yr7-10Nur Hidayah Abdul AzizPas encore d'évaluation
- The Marketing Advantages of Strong BrandsDocument26 pagesThe Marketing Advantages of Strong BrandsMadir MustaffaPas encore d'évaluation
- Ak Self MGMT SkillsDocument4 pagesAk Self MGMT SkillsHema LathaPas encore d'évaluation
- Resume Summary for Gaurav BhallaDocument2 pagesResume Summary for Gaurav BhallaGorav BhallaPas encore d'évaluation
- Enrolment Form Grade 12Document3 pagesEnrolment Form Grade 12Francis JavonilloPas encore d'évaluation
- Getting Started Learning Path overviewDocument1 pageGetting Started Learning Path overviewsimha1177Pas encore d'évaluation
- Cell Poposal Tempalte 1Document5 pagesCell Poposal Tempalte 1MAYANN CAPONPONPas encore d'évaluation
- Abhi New Front CorrectDocument5 pagesAbhi New Front CorrectVasu ThakurPas encore d'évaluation
- Website - ICQ FAQ DocumentDocument5 pagesWebsite - ICQ FAQ Documentamanda fengPas encore d'évaluation
- Parkour PDFDocument16 pagesParkour PDFbonengPas encore d'évaluation
- Dinesh Qa-Qc EngineerDocument6 pagesDinesh Qa-Qc EngineerDinesh SomwanshiPas encore d'évaluation
- Consumer PerceptionDocument32 pagesConsumer PerceptionYamini PaliwalPas encore d'évaluation
- English As A Foreign Language Teacher PDFDocument202 pagesEnglish As A Foreign Language Teacher PDFLidanPas encore d'évaluation
- CRM Implementation Case Study in Service CompanyDocument29 pagesCRM Implementation Case Study in Service CompanyKarrizzmatic0% (1)
- Scheme Guidelines for Industry Institute Partnership CellDocument28 pagesScheme Guidelines for Industry Institute Partnership CellhariselvarajPas encore d'évaluation
- Story stem narratives in clinical researchDocument8 pagesStory stem narratives in clinical researchCristina TatomirPas encore d'évaluation