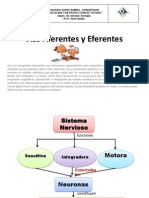
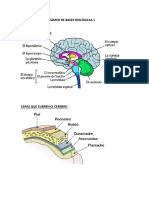
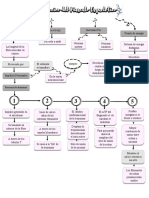
Vous aimerez peut-être aussi
- Biomecánica Del SaltoDocument20 pagesBiomecánica Del SaltoMiguel Puzzle Yermo100% (3)
- Cuestionarios Del Libro Ingeniería Industrial: Métodos, Estándares y Diseño Del TrabajoDocument19 pagesCuestionarios Del Libro Ingeniería Industrial: Métodos, Estándares y Diseño Del TrabajoErica Rodriguez100% (2)
- PRACTICA NO. 6-Receptores-Sensoriales EQ 5Document12 pagesPRACTICA NO. 6-Receptores-Sensoriales EQ 5Joguard Lofcraft100% (1)
- Laboratorio Electromiografí A UDocument16 pagesLaboratorio Electromiografí A UAlejandra BonillaPas encore d'évaluation
- Sistema Sensitivo MotorDocument1 pageSistema Sensitivo MotorBELISARIO LIMA SULLCAPas encore d'évaluation
- Humanismo y ConductismoDocument9 pagesHumanismo y ConductismoMerinyer Polanco100% (1)
- Funciones Corticales SuperioresDocument23 pagesFunciones Corticales SuperioresCarlos Rowe100% (5)
- Sensopercepcion 3er ParcialDocument3 pagesSensopercepcion 3er ParcialRoberto Carlos Mendez GalvanPas encore d'évaluation
- SISTEMA SENSITIVOMOTOR 2daDocument25 pagesSISTEMA SENSITIVOMOTOR 2daPaola Garcia MejiaPas encore d'évaluation
- Neurofisiologia Dolor PDFDocument27 pagesNeurofisiologia Dolor PDFkine_hectorPas encore d'évaluation
- Tallerneurociencia 120312070510 Phpapp01Document4 pagesTallerneurociencia 120312070510 Phpapp01Wilde Callata QuispePas encore d'évaluation
- Receptores y Efectores PDFDocument24 pagesReceptores y Efectores PDFcamiPas encore d'évaluation
- Guia N° 9 Receptores SensorialesDocument4 pagesGuia N° 9 Receptores Sensorialesʚiɞ Andre Bustamante Tagle ʚiɞPas encore d'évaluation
- 2565 BT 08 19 ReceptoresDocument24 pages2565 BT 08 19 ReceptoresRodrigoPas encore d'évaluation
- Exposición Sobre Receptores Sensitivos o Sensoriales o Sistema SensorialDocument9 pagesExposición Sobre Receptores Sensitivos o Sensoriales o Sistema SensorialLaura PaltaPas encore d'évaluation
- Fisiologia - CLASE - Somatosensorial 2017Document51 pagesFisiologia - CLASE - Somatosensorial 2017Stania CamposPas encore d'évaluation
- SENSIBILIDAD SOMÁTICA FisiologíaDocument15 pagesSENSIBILIDAD SOMÁTICA FisiologíaAyelen AbarzaPas encore d'évaluation
- Receptores Sensoriales, Circuitos Neuronales para Procesando Información PDFDocument81 pagesReceptores Sensoriales, Circuitos Neuronales para Procesando Información PDFItzel TejedaPas encore d'évaluation
- Semana 9 - Transducción Sensorial y Sensaciones SomáticasDocument40 pagesSemana 9 - Transducción Sensorial y Sensaciones SomáticasJaviera Lagos JaraPas encore d'évaluation
- Sensaciones Somaticas Transmision Vias SensitivasDocument72 pagesSensaciones Somaticas Transmision Vias SensitivasEvangelina JuradoPas encore d'évaluation
- Neurofisiologia DesarrolloDocument19 pagesNeurofisiologia DesarrolloMilagros M. PaucarPas encore d'évaluation
- Sensibilidad+General Receptores+CorporalesDocument71 pagesSensibilidad+General Receptores+CorporalesFlavio Méndez AparicioPas encore d'évaluation
- Receptor EsDocument11 pagesReceptor EsANDRES SALAZAR NIETOPas encore d'évaluation
- Unidad 3 Sistema Sensitivo y Motor. Equipo 5 Grupo 45686Document210 pagesUnidad 3 Sistema Sensitivo y Motor. Equipo 5 Grupo 45686BhanuDeviDasiPas encore d'évaluation
- Receptores ResumenDocument11 pagesReceptores ResumenNadia Alexandra Agurto100% (1)
- Sistema SensorialDocument22 pagesSistema SensorialHarold Ramos100% (1)
- Wuolah Free APUNTES T6Document18 pagesWuolah Free APUNTES T6Carolina R BPas encore d'évaluation
- Fisiologia de Los Receptores Nerviosos Perifericos (I)Document18 pagesFisiologia de Los Receptores Nerviosos Perifericos (I)Marco Almeida100% (2)
- Circuitos NeuronalesDocument8 pagesCircuitos NeuronalesEduardo Roque HernándezPas encore d'évaluation
- Los Receptores Sensoriales: ¿Qué Son?Document11 pagesLos Receptores Sensoriales: ¿Qué Son?SHAIDA P.EPas encore d'évaluation
- Sensopercepcion 170903212731Document134 pagesSensopercepcion 170903212731RUTH MARIA GARCIA HERNANDEZPas encore d'évaluation
- s.n.4 (Vias Aferentes y Eferentes)Document24 pagess.n.4 (Vias Aferentes y Eferentes)tornes9262Pas encore d'évaluation
- Psicologia de La PercepcionDocument57 pagesPsicologia de La Percepcionmererl14Pas encore d'évaluation
- Bloque 5 NerviosoDocument52 pagesBloque 5 NerviosoAnabel GutierrezPas encore d'évaluation
- Sistema SensorialDocument5 pagesSistema SensorialunhipodPas encore d'évaluation
- Sistema Sensorial Motor y de IntegraciónDocument14 pagesSistema Sensorial Motor y de IntegraciónErwin TrinidadPas encore d'évaluation
- Tema 7 Propiedades ReceptoresDocument5 pagesTema 7 Propiedades ReceptoresCarmeliaPas encore d'évaluation
- Tema 2Document6 pagesTema 2DAURA MEDINA HERNÁNDEZPas encore d'évaluation
- Tema 11Document18 pagesTema 11arancharr01Pas encore d'évaluation
- 1er Unidad, Atención y PDocument138 pages1er Unidad, Atención y PKaren Guisella Soledispa DiazPas encore d'évaluation
- Clase 4 Sentido AnimalDocument23 pagesClase 4 Sentido AnimalNicole TroncosoPas encore d'évaluation
- Sensación y PercepciónDocument32 pagesSensación y PercepciónAlexis VeraPas encore d'évaluation
- Funciones de Relación en Los Seres Vivos de La Madre TierraDocument10 pagesFunciones de Relación en Los Seres Vivos de La Madre TierrakmitagonzalesPas encore d'évaluation
- Receptor Sensorial - Wikipedia, La Enciclopedia LibreDocument27 pagesReceptor Sensorial - Wikipedia, La Enciclopedia LibreEmeron HoriangelizPas encore d'évaluation
- Resumen Neurobiología de Los Sentidos - Neurociencia IDocument15 pagesResumen Neurobiología de Los Sentidos - Neurociencia IAlexia Caroline50% (4)
- Temario Bases Biológicas 1Document25 pagesTemario Bases Biológicas 1Glendis Gonzalez SolisPas encore d'évaluation
- Sistema Nervioso Sensorial.: Aprendizaje Basado en ProblemasDocument38 pagesSistema Nervioso Sensorial.: Aprendizaje Basado en ProblemasAna Cristina AlvarezPas encore d'évaluation
- Cuestionario #7 .Document9 pagesCuestionario #7 .Ana TaverasPas encore d'évaluation
- Material Complementario ReceptoresDocument16 pagesMaterial Complementario ReceptoresAmarildo CristinoPas encore d'évaluation
- Somato SensorialDocument6 pagesSomato SensorialcandyPas encore d'évaluation
- Receptores SensorialesDocument10 pagesReceptores SensorialesJhuver Alex Campos VasquezPas encore d'évaluation
- Sensacion y PercepcionDocument40 pagesSensacion y PercepcionJaime HuenchunaoPas encore d'évaluation
- Funciones Sensoriales - Sistema SematosensorialDocument8 pagesFunciones Sensoriales - Sistema SematosensorialAndrea Bulnes MatamorosPas encore d'évaluation
- Modelo de Informe ChelitaDocument20 pagesModelo de Informe ChelitaSalomón ChávezPas encore d'évaluation
- Ps. Dra. Paula Díaz Vidiella CFT Tarapacá Arica-Chile 2020Document12 pagesPs. Dra. Paula Díaz Vidiella CFT Tarapacá Arica-Chile 2020Ana Isabel Castillo MuñozPas encore d'évaluation
- La Sensación PDFDocument19 pagesLa Sensación PDFmartaPas encore d'évaluation
- 14º Sesion para 4º Los Receptores Sensoriales y Actos ReflefosDocument18 pages14º Sesion para 4º Los Receptores Sensoriales y Actos ReflefosCasas C VictOrPas encore d'évaluation
- Los Receptores SensorialesDocument14 pagesLos Receptores SensorialesmixoneqPas encore d'évaluation
- Sistema Sensorial (Consolidado)Document34 pagesSistema Sensorial (Consolidado)Alx-sander MamaniPas encore d'évaluation
- Semana 1 Sistema SensorialDocument12 pagesSemana 1 Sistema SensorialLuis HerediaPas encore d'évaluation
- Receptores, EstimuloDocument2 pagesReceptores, EstimuloErik J. Roshan FloresPas encore d'évaluation
- Tema 11 PsicobioDocument17 pagesTema 11 PsicobioAlejandraPas encore d'évaluation
- Guia SensoperceptivaDocument6 pagesGuia SensoperceptivaAnsgabrielPas encore d'évaluation
- Trabajo de Fisio DisertacionDocument36 pagesTrabajo de Fisio Disertacionsusana_sanzanaPas encore d'évaluation
- Fisiología NeuromuscularDocument39 pagesFisiología NeuromuscularMaximiliano Di SantoPas encore d'évaluation
- Células y TejidosDocument47 pagesCélulas y TejidosKarlita MartínezPas encore d'évaluation
- CarneDocument69 pagesCarneMaria Paz Godoy100% (1)
- Historieta. Contracción MuscularDocument5 pagesHistorieta. Contracción MuscularCarlos MarguzPas encore d'évaluation
- MiofilamentosDocument17 pagesMiofilamentosJoshuaGaticaPas encore d'évaluation
- Clase No. 09 Union Neuromuscular Contracion MuscularDocument33 pagesClase No. 09 Union Neuromuscular Contracion MuscularMichael Jimenez MillonesPas encore d'évaluation
- Calidad Del Pescado Fresco FaoDocument299 pagesCalidad Del Pescado Fresco FaoRodolfito09100% (2)
- 001fisiologia Cardiovascular PUCDocument14 pages001fisiologia Cardiovascular PUCrodrincubusPas encore d'évaluation
- Tejido MuscularDocument36 pagesTejido MuscularBiología Celular E Histología Médica100% (3)
- Evaluacion Final TeoricaDocument11 pagesEvaluacion Final TeoricaLidia Lucila Marroquin JuarezPas encore d'évaluation
- Manual CTO 6ed - Cardiología y Cirugía CardiovascularDocument103 pagesManual CTO 6ed - Cardiología y Cirugía CardiovascularRaelina Gonzalez MoraPas encore d'évaluation
- Fisiologia Muscular y TetanoDocument6 pagesFisiologia Muscular y TetanoMia RodriguezPas encore d'évaluation
- Contraccion Muscular y Sistemas Energeticos - Lic. PoseDocument47 pagesContraccion Muscular y Sistemas Energeticos - Lic. PosePatricio Agustin VerónPas encore d'évaluation
- Control de Calidad de La Carne y Derivados CarnicosDocument61 pagesControl de Calidad de La Carne y Derivados CarnicosRenato Angulo Mamani50% (2)
- Mapa HistoDocument2 pagesMapa HistoKaroll Daniela PérezPas encore d'évaluation
- Guía Didáctica 3-EM PDFDocument36 pagesGuía Didáctica 3-EM PDFAndres CordobaPas encore d'évaluation
- Uniones Gap PDFDocument4 pagesUniones Gap PDFKristell Quezada100% (2)
- Tejido MuscularDocument12 pagesTejido MuscularEduardo GálvezPas encore d'évaluation
- Músculo LisoDocument13 pagesMúsculo Lisocepj1908Pas encore d'évaluation
- Sesión de Aprendizaje 18Document40 pagesSesión de Aprendizaje 18JHONATAN ACOSTA SEOPAPas encore d'évaluation
- Fisiologia Resumen de ClaseDocument148 pagesFisiologia Resumen de ClaseLevy McGardenPas encore d'évaluation
- Fisio Muscular de Las AvesDocument12 pagesFisio Muscular de Las AvesDiego Vasquez HuamaniPas encore d'évaluation
- Tejido Muscular 1 UES FMOCCDocument15 pagesTejido Muscular 1 UES FMOCCChristian LinaresPas encore d'évaluation
- La Química de Los Fenómenos CadavéricosDocument14 pagesLa Química de Los Fenómenos CadavéricosDANIELCAVERO16100% (1)
- UntitledDocument49 pagesUntitledandrea duranPas encore d'évaluation
- Tejido Muscular HistologíaDocument16 pagesTejido Muscular HistologíaAlberta SeguraPas encore d'évaluation