Vous aimerez peut-être aussi
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5794)
- Список обзоровDocument5 pagesСписок обзоровapi-3706027Pas encore d'évaluation
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (400)
- 18 Hartl TriggerDocument11 pages18 Hartl Triggerapi-3706027Pas encore d'évaluation
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- Article 15 PresentationDocument10 pagesArticle 15 Presentationapi-3706027Pas encore d'évaluation
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (895)
- 23 NacDocument7 pages23 Nacapi-3706027Pas encore d'évaluation
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- 19 SRPDocument6 pages19 SRPapi-3706027Pas encore d'évaluation
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- 20 DaviesDocument4 pages20 Daviesapi-3706027Pas encore d'évaluation
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- 22 BipDocument12 pages22 Bipapi-3706027Pas encore d'évaluation
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- 16 NFKBDocument10 pages16 NFKBapi-3706027Pas encore d'évaluation
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (588)
- 11 GapDocument10 pages11 Gapapi-3706027Pas encore d'évaluation
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (74)
- 12 tRNAmimicryDocument7 pages12 tRNAmimicryapi-3706027100% (1)
- 14 GFPDocument6 pages14 GFPapi-3706027Pas encore d'évaluation
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- 17 NicolaDocument5 pages17 Nicolaapi-3706027Pas encore d'évaluation
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (266)
- 09 TM RnaDocument6 pages09 TM Rnaapi-3706027Pas encore d'évaluation
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- 07 ChapevilleDocument7 pages07 Chapevilleapi-3706027Pas encore d'évaluation
- 02 Ogata NoharaDocument2 pages02 Ogata Noharaapi-3706027Pas encore d'évaluation
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- Synth Pro11Document1 pageSynth Pro11api-3706027Pas encore d'évaluation
- 04 CrickDocument6 pages04 Crickapi-3706027Pas encore d'évaluation
- 06 Brenner JacobDocument6 pages06 Brenner Jacobapi-3706027Pas encore d'évaluation
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2259)
- 05 Gros WatsonDocument5 pages05 Gros Watsonapi-3706027Pas encore d'évaluation
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- 01 SpirinDocument2 pages01 Spirinapi-3706027Pas encore d'évaluation
- Synth Pro11Document2 pagesSynth Pro11api-3706027Pas encore d'évaluation
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (344)
- Synth Pro11Document1 pageSynth Pro11api-3706027Pas encore d'évaluation
- Synth Pro11Document1 pageSynth Pro11api-3706027Pas encore d'évaluation
- Synth Pro10Document1 pageSynth Pro10api-3706027Pas encore d'évaluation
- Synth Pro10Document2 pagesSynth Pro10api-3706027Pas encore d'évaluation
- Synth Pro10Document1 pageSynth Pro10api-3706027Pas encore d'évaluation
- Synth Pro10Document1 pageSynth Pro10api-3706027Pas encore d'évaluation
- Synth Pro3Document3 pagesSynth Pro3api-3706027Pas encore d'évaluation
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (121)
- Synth Pro3Document2 pagesSynth Pro3api-3706027Pas encore d'évaluation
- 003 REVIEWER IN SCIENCE 7-SUgarDocument3 pages003 REVIEWER IN SCIENCE 7-SUgarrizza suarezPas encore d'évaluation
- WWW - Manabadi.co - in Timetables Svubsc07032009Document6 pagesWWW - Manabadi.co - in Timetables Svubsc07032009Lokesh ModemzPas encore d'évaluation
- 8th Class Biology Project WorkDocument10 pages8th Class Biology Project WorkPRANU83% (6)
- Water PotentialDocument35 pagesWater PotentialLuis GarcíaPas encore d'évaluation
- PNU Final Coaching Gen - EdDocument15 pagesPNU Final Coaching Gen - EdTatinPas encore d'évaluation
- CS 2000iDocument88 pagesCS 2000iBintang MaulanaPas encore d'évaluation
- SBRC HematologyOncology 2Document80 pagesSBRC HematologyOncology 2dalia khamoPas encore d'évaluation
- Health Status and Quality of Life of Geriatric Population in Old Age Homes and Living With Family in Chennai A Comparative StudyDocument5 pagesHealth Status and Quality of Life of Geriatric Population in Old Age Homes and Living With Family in Chennai A Comparative StudyInternational Journal of Innovative Science and Research TechnologyPas encore d'évaluation
- Ujmr 2020 - 4 - 24 - 30 PDFDocument7 pagesUjmr 2020 - 4 - 24 - 30 PDFUMYU Journal of Microbiology Research (UJMR)Pas encore d'évaluation
- Silviculture: February 2018Document69 pagesSilviculture: February 2018Tang YinPas encore d'évaluation
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- Induction Presentation Biology A Level and Gcse.204719226Document25 pagesInduction Presentation Biology A Level and Gcse.204719226Cut TirayaPas encore d'évaluation
- Applied Zoology: Pearl Culture Bechan LalDocument16 pagesApplied Zoology: Pearl Culture Bechan LalSk RajPas encore d'évaluation
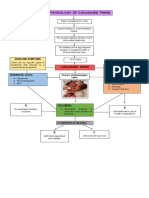
- Pathophysiology of Conjoined TwinsDocument1 pagePathophysiology of Conjoined TwinsPrincess Loreto AlegrePas encore d'évaluation
- Answers For Support Worksheet - Option F: 1 A Feature Bacteria Archaea EukaryotaDocument1 pageAnswers For Support Worksheet - Option F: 1 A Feature Bacteria Archaea EukaryotaHellö PëachPas encore d'évaluation
- 2457-Article Text-14907-2-10-20120724Document6 pages2457-Article Text-14907-2-10-20120724desi meleniaPas encore d'évaluation
- Cse Integration Lesson PlanDocument5 pagesCse Integration Lesson PlanCarl Anthony Lague PahuyoPas encore d'évaluation
- Strategies For Plant Regeneration From Mesophyll Protoplasts of The Recalcitrant Fruit and Farmwoodland Species L. (Sweet/wild Cherry)Document6 pagesStrategies For Plant Regeneration From Mesophyll Protoplasts of The Recalcitrant Fruit and Farmwoodland Species L. (Sweet/wild Cherry)Geral BlancoPas encore d'évaluation
- MFK-Drug Management CycleDocument32 pagesMFK-Drug Management CycleLhynda Priarti Latif100% (1)
- Anima and Animus Term ReportDocument17 pagesAnima and Animus Term ReportSadia Shahid100% (1)
- Unit 3Document61 pagesUnit 3JessicaHaePas encore d'évaluation
- Characteristics of A Community PDFDocument15 pagesCharacteristics of A Community PDFAlijanPas encore d'évaluation
- Lesson 3.1 History of Life On EarthDocument1 pageLesson 3.1 History of Life On EarthJoshua EboraPas encore d'évaluation
- Lactic Acid Production From Cane MolassesDocument4 pagesLactic Acid Production From Cane MolassesSenkatuuka LukePas encore d'évaluation
- Hubungan Tingkat Pengetahuan Dan Dukungan Keluarga Terhadap Kepatuhan Ibu Hamil Mengkonsumsi Tablet Zat BesiDocument13 pagesHubungan Tingkat Pengetahuan Dan Dukungan Keluarga Terhadap Kepatuhan Ibu Hamil Mengkonsumsi Tablet Zat Besinovilia eka sariPas encore d'évaluation
- Upcoming Predicted Material For PTE-A Exam Feb-2019Document123 pagesUpcoming Predicted Material For PTE-A Exam Feb-2019GauravPas encore d'évaluation
- Aspb 2012 - 106Document204 pagesAspb 2012 - 106arunprabhu_dhanapalPas encore d'évaluation
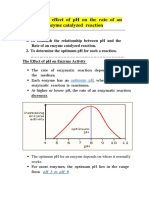
- Effect of PH On Enzyme ActivityDocument12 pagesEffect of PH On Enzyme ActivityAb AbPas encore d'évaluation
- Treatment of Latent Tuberculosis InfectionDocument10 pagesTreatment of Latent Tuberculosis InfectionRichard WoolitePas encore d'évaluation
- A Brief Introduction To ArgumentsDocument7 pagesA Brief Introduction To ArgumentsJohn DarwinPas encore d'évaluation
- (English (Auto-Generated) ) Ep. 94 - Andres Ryan of BioAquatix - Com On Breeding Clown Loaches (DownSub - Com)Document78 pages(English (Auto-Generated) ) Ep. 94 - Andres Ryan of BioAquatix - Com On Breeding Clown Loaches (DownSub - Com)acromontiPas encore d'évaluation