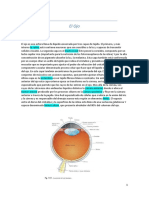
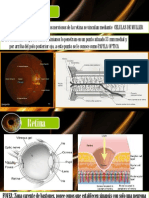
Vous aimerez peut-être aussi
- Cuadro Comparativo TCC y TRECDocument3 pagesCuadro Comparativo TCC y TRECSalvador Llenas Remigio75% (4)
- Neuro VisionDocument4 pagesNeuro VisionSol SilvaPas encore d'évaluation
- Fotoquimica de La VisionDocument8 pagesFotoquimica de La VisionMiguel Alfredo Ochoa Ipushima100% (1)
- Descripcion de Curso de TrhaDocument10 pagesDescripcion de Curso de Trharotoplas_001Pas encore d'évaluation
- Bloques Funcionales Cerebra LesDocument6 pagesBloques Funcionales Cerebra LesRafael Eduardo Pesantes Roncal100% (2)
- Imforme Biofisica de La VisiónDocument5 pagesImforme Biofisica de La Visiónpurple skyPas encore d'évaluation
- Ciclo visual: procesamiento de la información lumínicaDocument27 pagesCiclo visual: procesamiento de la información lumínicaHsieh Lo MingPas encore d'évaluation
- Taller Transducción VisualDocument3 pagesTaller Transducción VisualJoice LadinoPas encore d'évaluation
- FotorreceptoresDocument2 pagesFotorreceptoresAnonymous HQuX2LUsDPas encore d'évaluation
- Word REtinaDocument89 pagesWord REtinaCesar Alex Lumbre YuptonPas encore d'évaluation
- Sistema Central Visual CompletoDocument17 pagesSistema Central Visual CompletoLily Julieth Avila PrietoPas encore d'évaluation
- La transducción visual: del estímulo a la percepciónDocument2 pagesLa transducción visual: del estímulo a la percepciónMelissa MatutePas encore d'évaluation
- FISIOLOGIA DE LA VISION Y REFLEJOS PUPILARESDocument41 pagesFISIOLOGIA DE LA VISION Y REFLEJOS PUPILARESOscar Padilla Navarro100% (1)
- Túnica Nerviosa o Retina 11Document34 pagesTúnica Nerviosa o Retina 11henzejuly50% (2)
- Potenciales Evocados VisualesDocument11 pagesPotenciales Evocados VisualesCa SalPas encore d'évaluation
- VisiónDocument17 pagesVisiónDavidPas encore d'évaluation
- RetinaDocument2 pagesRetinaa318937Pas encore d'évaluation
- Clase 4 Vision-Clase-NeurofisiologiaDocument31 pagesClase 4 Vision-Clase-Neurofisiologialuis HenriquePas encore d'évaluation
- Capas de La RetinaDocument5 pagesCapas de La RetinaOrlinton Noé Rosales RodríguezPas encore d'évaluation
- Trabajo de Fisiología-2Document5 pagesTrabajo de Fisiología-2Mafer BarrazaPas encore d'évaluation
- Fisiología visiónDocument33 pagesFisiología visiónJuan Carlos Monte GonzálezPas encore d'évaluation
- Presentación Fisiología SensorialDocument21 pagesPresentación Fisiología Sensorialelizabeth lazoPas encore d'évaluation
- La RetinaDocument25 pagesLa RetinaOpticos Services OpticaPas encore d'évaluation
- La RetinaDocument17 pagesLa RetinaEderth Cordero100% (1)
- Recorrido de La VisiónDocument7 pagesRecorrido de La VisiónAleja BeltranPas encore d'évaluation
- Extra 3. Sensibilidad VisualDocument7 pagesExtra 3. Sensibilidad VisualAlbaPas encore d'évaluation
- Tarea 3 - Via Visual PDFDocument30 pagesTarea 3 - Via Visual PDFJaimeSergioVasquezZunigaPas encore d'évaluation
- Fisiologia Sensorial de La Via OpticaDocument45 pagesFisiologia Sensorial de La Via OpticaGabrielaPas encore d'évaluation
- Mecanismo de transducción visualDocument38 pagesMecanismo de transducción visualChuy Cervantes AguilarPas encore d'évaluation
- Anatomía, Histología y Fisiología RetinalDocument24 pagesAnatomía, Histología y Fisiología RetinalJuan Carlos NovoaPas encore d'évaluation
- Unidad 2 VisiónDocument8 pagesUnidad 2 VisiónSol AlderetePas encore d'évaluation
- Fototransducción VisualDocument7 pagesFototransducción VisualCynthia Jaramillo MachucaPas encore d'évaluation
- Retina OtraDocument8 pagesRetina OtraJm GamaPas encore d'évaluation
- Tema 3 Función VisualDocument6 pagesTema 3 Función VisualMaria GarciaPas encore d'évaluation
- Retina en EsDocument25 pagesRetina en EsJorge Leonardo BedoyaPas encore d'évaluation
- Fototransduccion VisualDocument7 pagesFototransduccion VisualAnonymous HQuX2LUsDPas encore d'évaluation
- Tema 4. Psicobiología de La Visión. Versión DefinitivaDocument10 pagesTema 4. Psicobiología de La Visión. Versión DefinitivaManuel Cañas LucendoPas encore d'évaluation
- Sensacion y PercepcionDocument16 pagesSensacion y PercepcionEstephany Avila ArenasPas encore d'évaluation
- Anatomía MicroscópicaDocument8 pagesAnatomía MicroscópicaConejita MorenoPas encore d'évaluation
- Función receptora y nerviosa de la retinaDocument10 pagesFunción receptora y nerviosa de la retinaJhony PradelaPas encore d'évaluation
- wuolah-free-TEMA 1 - Maduración RetinaDocument10 pageswuolah-free-TEMA 1 - Maduración RetinaevePas encore d'évaluation
- ElectrorretinogramaDocument12 pagesElectrorretinogramaClaudio Esteban Osses RiverosPas encore d'évaluation
- Tesis BCadiz 2020 v4 PDFDocument78 pagesTesis BCadiz 2020 v4 PDFDaniel GonzálezPas encore d'évaluation
- Sistema Visual (Resumen)Document3 pagesSistema Visual (Resumen)Rosendblack100% (1)
- Paper Eog Con IntroDocument12 pagesPaper Eog Con IntroNicolas Medina ChiloPas encore d'évaluation
- Apuntes oftalmología UVDocument158 pagesApuntes oftalmología UVSebastian MontenegroPas encore d'évaluation
- Avance PaperDocument6 pagesAvance PaperMayki de la CruzPas encore d'évaluation
- Taller Fotoquimica de La VisionDocument6 pagesTaller Fotoquimica de La VisionGladys Peláez100% (1)
- Fisiologia VisionDocument16 pagesFisiologia VisionAnonymous g8P3dNxPas encore d'évaluation
- Informe Sobre La Fisiologia de La VisionDocument3 pagesInforme Sobre La Fisiologia de La VisionDiana HernandezPas encore d'évaluation
- Histologia de OjoDocument20 pagesHistologia de OjoKevin Mercado GarciaPas encore d'évaluation
- Transduccion VisualDocument14 pagesTransduccion VisualErika Llanos Mora50% (2)
- Informe OjoDocument4 pagesInforme OjoandreaPas encore d'évaluation
- RETINADocument9 pagesRETINAKARLA CELPas encore d'évaluation
- Alumbrado Practica 2Document16 pagesAlumbrado Practica 2Flores Gonzales Marcela LauraPas encore d'évaluation
- Importancia LCPUFA visiónDocument11 pagesImportancia LCPUFA visiónJose Muñoz100% (1)
- Anatomia y Fisiologia OjoDocument38 pagesAnatomia y Fisiologia OjoMariana TakataPas encore d'évaluation
- Capitulo 12 Ganong VistaDocument46 pagesCapitulo 12 Ganong VistaFernando PortilloPas encore d'évaluation
- Anatomía de la vía visual humanaDocument9 pagesAnatomía de la vía visual humanaMaria Jose MolinaPas encore d'évaluation
- Resumen de Sistema Visual Página 267 PDFDocument3 pagesResumen de Sistema Visual Página 267 PDFCesibel RamirezPas encore d'évaluation
- Pigmentos visuales y células de la retina enDocument20 pagesPigmentos visuales y células de la retina enKevin VallesPas encore d'évaluation
- Fisiología de La RetinaDocument24 pagesFisiología de La RetinaozkrefrainPas encore d'évaluation
- El cannabis en patologías del sistema nervioso centralD'EverandEl cannabis en patologías del sistema nervioso centralPas encore d'évaluation
- Francisca No SDocument13 pagesFrancisca No SAlex Goldschmidt BozzoPas encore d'évaluation
- 2 8 SistemaEndocrinoDocument18 pages2 8 SistemaEndocrinoppPas encore d'évaluation
- Tecnicas para Evitar ColisionesDocument22 pagesTecnicas para Evitar ColisionesWilliam RoaPas encore d'évaluation
- Evaluacion Final Mod 5Document5 pagesEvaluacion Final Mod 5ppPas encore d'évaluation
- I86 TutorialDocument3 pagesI86 TutorialppPas encore d'évaluation
- PsicolingüísticaDocument8 pagesPsicolingüísticaEdwin Mauricio Lopez BallesterosPas encore d'évaluation
- Fase 4 - Teoria ConstructivistaDocument9 pagesFase 4 - Teoria Constructivistaana teheran100% (1)
- 2 Obtención de Valores Normativos en La Evaluación de La Función Auditiva CentralDocument5 pages2 Obtención de Valores Normativos en La Evaluación de La Función Auditiva CentralFrancisca Aravena RoaPas encore d'évaluation
- El Entrenamiento MentalDocument8 pagesEl Entrenamiento MentalMarko Antonio SPas encore d'évaluation
- Urgencias Subjetivas Un Modo de Psicoanalisis Aplicado UltimoDocument67 pagesUrgencias Subjetivas Un Modo de Psicoanalisis Aplicado UltimoDiego BilbaoPas encore d'évaluation
- Como Aprende El CerebroDocument12 pagesComo Aprende El Cerebrogil2Pas encore d'évaluation
- Tabla Comparativa de TeoriasDocument2 pagesTabla Comparativa de TeoriasYudith OlivarezPas encore d'évaluation
- Sesgos CognitivosDocument10 pagesSesgos CognitivosAlejandro CallePas encore d'évaluation
- Programa Educativo Integra1 Ficha de RegistroDocument128 pagesPrograma Educativo Integra1 Ficha de RegistroRocio Olivera DiazPas encore d'évaluation
- Tecnica de EntrevistaDocument2 pagesTecnica de EntrevistaYarleny NolascoPas encore d'évaluation
- Ficha # 5 Lengua 2 BguDocument3 pagesFicha # 5 Lengua 2 Bgucgranizo827Pas encore d'évaluation
- AutismoDocument3 pagesAutismoGerardo AragónPas encore d'évaluation
- Rewire Your Anxious Brain - How To Use The Neuroscience of Fear To End Anxiety, Panic, and Worry (PDFDrive)Document234 pagesRewire Your Anxious Brain - How To Use The Neuroscience of Fear To End Anxiety, Panic, and Worry (PDFDrive)Liana Pérez RodríguezPas encore d'évaluation
- Resumen de La Historia de La NeuropsicologiaDocument1 pageResumen de La Historia de La Neuropsicologiaprincesslzt100% (9)
- Violeta Tapia Y Jorge Luna - Procesos Cognitivos y Desempeño LectorDocument32 pagesVioleta Tapia Y Jorge Luna - Procesos Cognitivos y Desempeño LectorVAHRPas encore d'évaluation
- Habilidades psicomotrices y pensamientoDocument2 pagesHabilidades psicomotrices y pensamientoAlfonso Cuetto BornacellyPas encore d'évaluation
- Sihara Calderin-InformeDocument1 pageSihara Calderin-InformeÁlvaro Javier Sánchez BeltránPas encore d'évaluation
- Crecimiento y Desarrollo SdajDocument4 pagesCrecimiento y Desarrollo Sdajmichelle100% (1)
- Resumen Cap. 2 Materia y Memoria, BergsonDocument10 pagesResumen Cap. 2 Materia y Memoria, Bergsonteresita100% (1)
- Corteza PrefrontalDocument9 pagesCorteza PrefrontalIris LópezPas encore d'évaluation
- Compendio de Psicología: definición, objetivos y métodosDocument100 pagesCompendio de Psicología: definición, objetivos y métodosJosué Rodrigo RVPas encore d'évaluation
- Teoría de La Personalidad Tarea 4Document6 pagesTeoría de La Personalidad Tarea 4Mariel Valdez100% (1)
- Estrés en estudiantes de psicología de la UNIMINUTO en MocoaDocument29 pagesEstrés en estudiantes de psicología de la UNIMINUTO en MocoaMILLERPas encore d'évaluation
- La PsicoterapiaDocument11 pagesLa PsicoterapiaAlexandra AlvizuPas encore d'évaluation
- Psicobiología BUPADocument7 pagesPsicobiología BUPAElseys RuedaPas encore d'évaluation
- Inventario de Perfil EstructuralDocument2 pagesInventario de Perfil EstructuralSarita MartinezPas encore d'évaluation
- Elementos de PsicologíaDocument9 pagesElementos de PsicologíaEmanuel MoreyraPas encore d'évaluation