Vous aimerez peut-être aussi
- Brothers: Integration of Metabolism and Metabolic Processes in Fed, Fasting and Starvation StatesDocument14 pagesBrothers: Integration of Metabolism and Metabolic Processes in Fed, Fasting and Starvation StatesNischal DhPas encore d'évaluation
- Lecture-5 Hormonal Regulation of Intestinal MotilityDocument4 pagesLecture-5 Hormonal Regulation of Intestinal Motilityمرتضى حسين عبدPas encore d'évaluation
- CH11-Amino Acid MetabolismDocument106 pagesCH11-Amino Acid MetabolismChatchawinPas encore d'évaluation
- Report EnzymesDocument34 pagesReport Enzymesnestie villavirayPas encore d'évaluation
- 16 Pathology of Lipid MetDocument52 pages16 Pathology of Lipid MetyasingadaPas encore d'évaluation
- Biochem Lec Term Paper HypoglycemiaDocument8 pagesBiochem Lec Term Paper Hypoglycemiaapi-318284296Pas encore d'évaluation
- Grape Seed Extract (Vitis Vinifera) Partially Reverses High Fat Diet-Induced Obesity in C57BL/6J MiceDocument7 pagesGrape Seed Extract (Vitis Vinifera) Partially Reverses High Fat Diet-Induced Obesity in C57BL/6J MiceMohammad SutamiPas encore d'évaluation
- Lipid and Glucose Methodology PDFDocument16 pagesLipid and Glucose Methodology PDFJezzah Mae CañetePas encore d'évaluation
- Tryptophan Metabolism: Dr. Ashok Kumar J International Medical School Management and Science University MalaysiaDocument23 pagesTryptophan Metabolism: Dr. Ashok Kumar J International Medical School Management and Science University MalaysiaShah HuzaifaPas encore d'évaluation
- SDLS 2008 Serum Total Protein and Albumin DeterminationDocument2 pagesSDLS 2008 Serum Total Protein and Albumin DeterminationgenieqtPas encore d'évaluation
- Glycoproteins: Charlotte V. Bañes, MDDocument71 pagesGlycoproteins: Charlotte V. Bañes, MDswitimahalePas encore d'évaluation
- Biochemistry: Ihrevale LIPID META Bolism 1Document92 pagesBiochemistry: Ihrevale LIPID META Bolism 1Roy BelenPas encore d'évaluation
- A Brief Overview of Hemoglobin ElectrophoresisDocument46 pagesA Brief Overview of Hemoglobin Electrophoresisمجاهد إسماعيل حسن حسينPas encore d'évaluation
- Body FluidsDocument24 pagesBody FluidsMohamed MidoPas encore d'évaluation
- B.D.S Course Biochemistry/Syllabus: (With Effect From 2010-11 Onwards)Document4 pagesB.D.S Course Biochemistry/Syllabus: (With Effect From 2010-11 Onwards)AnandPas encore d'évaluation
- Introduction To Biochemistry: by Zaheer Uddin (Pharm-D, M Phil (Pharmacy Practice) MBA (MARKETING)Document63 pagesIntroduction To Biochemistry: by Zaheer Uddin (Pharm-D, M Phil (Pharmacy Practice) MBA (MARKETING)Izat KhanPas encore d'évaluation
- Concise Biochemistry: Fundamental Principles: March 2016Document52 pagesConcise Biochemistry: Fundamental Principles: March 2016Sagar DeshmaniPas encore d'évaluation
- 2 - Lipid Metabolism Lecture For StudentsDocument68 pages2 - Lipid Metabolism Lecture For StudentshwhsgxPas encore d'évaluation
- Written Exam Blue PrintDocument12 pagesWritten Exam Blue PrintMotasim_mPas encore d'évaluation
- Principles of Dna Isolation & PurificationDocument10 pagesPrinciples of Dna Isolation & PurificationRatish Singh50% (2)
- First Professional Mbbs Fourth Semester EXAMINATION SESSION-2007-2008 Subject - BiochemistryDocument9 pagesFirst Professional Mbbs Fourth Semester EXAMINATION SESSION-2007-2008 Subject - BiochemistryIsrasami NaparPas encore d'évaluation
- General Introduction and Basic Biochemical PrinciplesDocument6 pagesGeneral Introduction and Basic Biochemical PrinciplescelecosibPas encore d'évaluation
- Git Biochemistry of GitDocument9 pagesGit Biochemistry of GitSibatPas encore d'évaluation
- Sphingolipids: Dr. Abir Alghanouchi Biochemistry Department Sciences CollegeDocument12 pagesSphingolipids: Dr. Abir Alghanouchi Biochemistry Department Sciences CollegeUzac BenuPas encore d'évaluation
- Complex Carbohydrates - GlycoproteinsDocument28 pagesComplex Carbohydrates - GlycoproteinsJessica SannohPas encore d'évaluation
- Diabetes Mellitus: Overview and TreatmentsDocument36 pagesDiabetes Mellitus: Overview and TreatmentsGlenn Nadine N. Masicampo100% (1)
- Molecular CloningDocument61 pagesMolecular CloningDavi DzikirianPas encore d'évaluation
- Creatine and Creatinine MetabolismDocument108 pagesCreatine and Creatinine MetabolismMae Matira AbeladorPas encore d'évaluation
- Cholesterol - Synthesis, Metabolism, Regulation PDFDocument10 pagesCholesterol - Synthesis, Metabolism, Regulation PDFAdreiTheTripleA100% (1)
- Protein SynthesisDocument18 pagesProtein Synthesisabisantiago6131100% (1)
- 19.7 Membrane Lipids: Phospholipids: Group 3Document36 pages19.7 Membrane Lipids: Phospholipids: Group 3Masbateyeandlaser EyeclinicPas encore d'évaluation
- ChromatographyDocument31 pagesChromatographyarun231187Pas encore d'évaluation
- Metabolism of Carbohydrate: Department of Biochemistry Faculty of Medicine University of YARSI JakartaDocument60 pagesMetabolism of Carbohydrate: Department of Biochemistry Faculty of Medicine University of YARSI JakartaAmanda PutriPas encore d'évaluation
- Hexose Monophosphate ShuntDocument43 pagesHexose Monophosphate ShuntSecret AgentPas encore d'évaluation
- 3dr Year Common Biochemistry MCQsDocument8 pages3dr Year Common Biochemistry MCQsYasmin Amr NounouPas encore d'évaluation
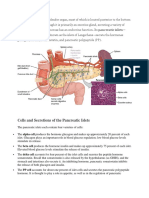
- Cells and Secretions of The Pancreatic IsletsDocument4 pagesCells and Secretions of The Pancreatic IsletsSophia OcayPas encore d'évaluation
- LipidsDocument13 pagesLipidsalianaPas encore d'évaluation
- Biochemistry of GlycoproteinDocument6 pagesBiochemistry of GlycoproteinMahathir Mohmed100% (7)
- s15 Miller Chap 3b LectureDocument25 pagess15 Miller Chap 3b LectureDorice Clement100% (1)
- Fatty Acid Synthesis 11.12.19Document18 pagesFatty Acid Synthesis 11.12.19Sanreet Randhawa100% (1)
- Biochemistry - C4 Proteins Determination of Primary StructureDocument2 pagesBiochemistry - C4 Proteins Determination of Primary StructureKim LlamasPas encore d'évaluation
- Bioactive PeptidesDocument16 pagesBioactive PeptidesOmotoyinbo SegunPas encore d'évaluation
- Purine & Pyrimidine MetabolismDocument22 pagesPurine & Pyrimidine MetabolismManda100% (1)
- MCQ MembranesDocument10 pagesMCQ MembranesMarilyne RizkPas encore d'évaluation
- B Y: - Idr. Megha Gaur BDS IDocument89 pagesB Y: - Idr. Megha Gaur BDS IRishab GaurPas encore d'évaluation
- Glucose, Part1Document33 pagesGlucose, Part1SarahPas encore d'évaluation
- Abnormal Constituents of UrineDocument2 pagesAbnormal Constituents of UrineAries DocPas encore d'évaluation
- Digestive System: Digestion and Absorption of Carbohydrates, Proteins and FatsDocument45 pagesDigestive System: Digestion and Absorption of Carbohydrates, Proteins and FatshiPas encore d'évaluation
- 41 & 42 - Nucleic Acid MetabolismDocument56 pages41 & 42 - Nucleic Acid MetabolismMădă Claws100% (1)
- HMPDocument44 pagesHMPraanja2Pas encore d'évaluation
- Chemistryofproteinswithclinicalapplications 190621192525 PDFDocument181 pagesChemistryofproteinswithclinicalapplications 190621192525 PDFAl-waleed Julkanain100% (1)
- Cardiac MetabolismDocument35 pagesCardiac Metabolismwaqas_xsPas encore d'évaluation
- Hormones and Metabolic RegulationDocument40 pagesHormones and Metabolic RegulationEka HabinaPas encore d'évaluation
- Biochem - (With Answers and Achievement Chart)Document11 pagesBiochem - (With Answers and Achievement Chart)Valine Cysteine MethioninePas encore d'évaluation
- Practical BiochemistryDocument35 pagesPractical BiochemistryMockinjay100% (1)
- Format of Animal Development Practical Report:: Text Book Berbahasa Indonesia Atau Bahasa InggrisDocument14 pagesFormat of Animal Development Practical Report:: Text Book Berbahasa Indonesia Atau Bahasa InggrisSafira Dwi OktavianiPas encore d'évaluation
- Citric Acid Cycle - Pyruvate DehydrogenaseDocument33 pagesCitric Acid Cycle - Pyruvate Dehydrogenasesultan khabeeb100% (1)
- Ion TransportD'EverandIon TransportDavid KeelingPas encore d'évaluation
- Estella Carganilla - Presentation For Chaffee Zoo Animal 2018Document35 pagesEstella Carganilla - Presentation For Chaffee Zoo Animal 2018api-395970440Pas encore d'évaluation
- M220008 Euthaphen Product InsertDocument1 pageM220008 Euthaphen Product InsertnadirhussainbuzdarPas encore d'évaluation
- 1 Eye of Horus and Count Like An Egyptian 2Document6 pages1 Eye of Horus and Count Like An Egyptian 2yin yinPas encore d'évaluation
- Chapter 22 Maternal NotesDocument2 pagesChapter 22 Maternal NotesChin T. OndongPas encore d'évaluation
- Sample Paper 1 ENGLISHDocument22 pagesSample Paper 1 ENGLISHDhawan ChoudharyPas encore d'évaluation
- Jane Goodall: Navigation SearchDocument8 pagesJane Goodall: Navigation SearchVeena HegdePas encore d'évaluation
- NEBDN Dental Charting Book APRIL 2015Document13 pagesNEBDN Dental Charting Book APRIL 2015Avita RathPas encore d'évaluation
- Pedia BulletsDocument2 pagesPedia BulletsDebbiearlPas encore d'évaluation
- Minimal Pairs Pronunciation PracticeDocument2 pagesMinimal Pairs Pronunciation Practiceadrian ariasPas encore d'évaluation
- DanMachi Volume 05 - Chapter 1Document32 pagesDanMachi Volume 05 - Chapter 1Joe ForsaPas encore d'évaluation
- Sexout ModsDocument7 pagesSexout ModsPablo Huerdo100% (2)
- Harga Baru Nestle OkeDocument8 pagesHarga Baru Nestle OkeFitri 'cwett' WijayantiPas encore d'évaluation
- Schleich Knights MagazineDocument16 pagesSchleich Knights Magazinebananica28100% (1)
- Vitamins and Minerals: Nutrient (Vitamins) Needed For Key SourcesDocument4 pagesVitamins and Minerals: Nutrient (Vitamins) Needed For Key SourcesKevin Carl A. CorpuzPas encore d'évaluation
- PublicationDocument56 pagesPublicationAlvarin CsboyPas encore d'évaluation
- BeefDocument12 pagesBeefTheamicable Firstlady MarPas encore d'évaluation
- Algorithms For IV Fluid Therapy in Children and Young People in Hospital Set of 6 PDF 2190274957 PDFDocument6 pagesAlgorithms For IV Fluid Therapy in Children and Young People in Hospital Set of 6 PDF 2190274957 PDFFurqon AfandriPas encore d'évaluation
- Unit 2: Academic ReadingDocument5 pagesUnit 2: Academic ReadingJuliana XavierPas encore d'évaluation
- Major and Minor ConnectorsDocument6 pagesMajor and Minor ConnectorsAntal Barbela67% (3)
- Macros HunterDocument5 pagesMacros HunterikenconstructPas encore d'évaluation
- Connective TissueDocument21 pagesConnective TissueCalvin Lin Jia RongPas encore d'évaluation
- Report Text AboutDocument3 pagesReport Text Aboutawe riantoPas encore d'évaluation
- Zoology BiomesDocument11 pagesZoology BiomesDennie Zody LoganPas encore d'évaluation
- Hematology Basics NewDocument32 pagesHematology Basics Newkimbo23Pas encore d'évaluation
- RFTPS2CULTDocument7 pagesRFTPS2CULTAna Vaquer MontoroPas encore d'évaluation
- Narrative TextDocument5 pagesNarrative TextWinniePas encore d'évaluation
- Uas GenapDocument10 pagesUas GenapDream32apPas encore d'évaluation
- EpisiotomyDocument2 pagesEpisiotomydanur kusuma arini putriPas encore d'évaluation
- Kode TopografiDocument69 pagesKode TopografiMUHAMMAD RASYID RPas encore d'évaluation
- 上海市虹口区2017学年度第一学期期中考试七年级英语学科试卷(PDF版 答案)Document5 pages上海市虹口区2017学年度第一学期期中考试七年级英语学科试卷(PDF版 答案)Lidia HuangPas encore d'évaluation