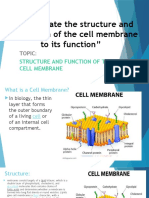
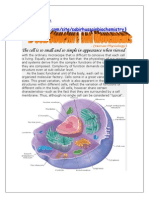
Vous aimerez peut-être aussi
- Cell Membrane: Navigation SearchDocument10 pagesCell Membrane: Navigation SearchMuhassanah Abd AzizPas encore d'évaluation
- Cell Notes Prokaryotic Cell, CM, CWDocument7 pagesCell Notes Prokaryotic Cell, CM, CWSaumya GopalPas encore d'évaluation
- 2structure and Functions of Cell MembraneDocument19 pages2structure and Functions of Cell MembraneedriansamaPas encore d'évaluation
- Cell Membranes and Transport As BiologyDocument41 pagesCell Membranes and Transport As BiologyADEEL AHMADPas encore d'évaluation
- SF - MD.0145.T.22 Ally Omary MwambelaDocument10 pagesSF - MD.0145.T.22 Ally Omary MwambelaAlly OmaryPas encore d'évaluation
- Chapter 2 Student Copy ZoologyDocument5 pagesChapter 2 Student Copy Zoologyapi-530699294Pas encore d'évaluation
- Full Download Zoology 9th Edition Miller Solutions ManualDocument35 pagesFull Download Zoology 9th Edition Miller Solutions Manualcaveneywilliams100% (14)
- Biomembrane: Selective PermeabilityDocument2 pagesBiomembrane: Selective PermeabilityKrystel LahomPas encore d'évaluation
- $RD2KRQ4Document32 pages$RD2KRQ4imtiazakhtar012Pas encore d'évaluation
- Cell MembraneDocument15 pagesCell MembraneAyesha SaleemPas encore d'évaluation
- Functional and Structural Properties of Natural BiomembranesDocument7 pagesFunctional and Structural Properties of Natural BiomembranesAhsan RazaPas encore d'évaluation
- Cell MembranesDocument5 pagesCell MembranesJANET ANNE BORROMEOPas encore d'évaluation
- Chapter 2 Student Copy ZoologyDocument5 pagesChapter 2 Student Copy Zoologyapi-523871804Pas encore d'évaluation
- Official Chapter 2 Student Copy ZoologyDocument5 pagesOfficial Chapter 2 Student Copy Zoologyapi-528700386Pas encore d'évaluation
- LO Week 7Document22 pagesLO Week 7Matthew ChristopherPas encore d'évaluation
- CMT 2021-22 by Mr. Adeel AhmadDocument24 pagesCMT 2021-22 by Mr. Adeel AhmadADEEL AHMADPas encore d'évaluation
- Cells: Transport of Materials Across The Cell MembraneDocument6 pagesCells: Transport of Materials Across The Cell MembraneSara SalmanPas encore d'évaluation
- Cell MembraneDocument10 pagesCell Membraneharshika tembhurnePas encore d'évaluation
- Cell Structure & MembraneDocument24 pagesCell Structure & MembraneAbhishek Singh ChandelPas encore d'évaluation
- Cell MembraneDocument29 pagesCell Membranesyraregencia15Pas encore d'évaluation
- Morphological Function of The Cell Presentation PHDocument52 pagesMorphological Function of The Cell Presentation PHDoc HamsPas encore d'évaluation
- Unit 2 ReviewerDocument4 pagesUnit 2 Revieweraaleah moscaPas encore d'évaluation
- 02 Cell PhysiologyDocument28 pages02 Cell PhysiologyZaw Lin TheinPas encore d'évaluation
- Cell Structure: Prepared by Dr. Sana AslamDocument41 pagesCell Structure: Prepared by Dr. Sana Aslamsajjad Ali balouch100% (1)
- Prokaryotic and Eukaryotic CellsDocument6 pagesProkaryotic and Eukaryotic CellsJam MoralejaPas encore d'évaluation
- Cell Biology and Membrane BiochemistryDocument106 pagesCell Biology and Membrane BiochemistryVijay Bhasker TekulapallyPas encore d'évaluation
- Lecture 3 Notes Cellular Basis of LifeDocument11 pagesLecture 3 Notes Cellular Basis of Lifeochienganthony001Pas encore d'évaluation
- Animal Physiology Theory Lec1Document24 pagesAnimal Physiology Theory Lec1ao868598Pas encore d'évaluation
- Cell StructureDocument10 pagesCell StructureJane NadalPas encore d'évaluation
- Cell Membrane Transport ActivityDocument7 pagesCell Membrane Transport ActivityoctavianojaninePas encore d'évaluation
- Bio Final AssignmentDocument47 pagesBio Final AssignmentmikhailPas encore d'évaluation
- Module 10 ACTDocument5 pagesModule 10 ACTLeighPas encore d'évaluation
- Screenshot 2023-09-26 at 8.50.46 PMDocument40 pagesScreenshot 2023-09-26 at 8.50.46 PMnomysalihPas encore d'évaluation
- Cell Structure by Sabir.h.nDocument15 pagesCell Structure by Sabir.h.nprinceaimy.bestPas encore d'évaluation
- 8 Cell NotesDocument7 pages8 Cell NotesAbhijeet AmetaPas encore d'évaluation
- Cell Membrane: Dr. Samia ShahbazDocument26 pagesCell Membrane: Dr. Samia Shahbazreema aslamPas encore d'évaluation
- Genzoo Module 1 Quiz Page 2 13Document12 pagesGenzoo Module 1 Quiz Page 2 13Joan Mae YaoPas encore d'évaluation
- Full Download Human Anatomy 4th Edition Saladin Solutions ManualDocument35 pagesFull Download Human Anatomy 4th Edition Saladin Solutions Manualvocality.duettoqw603100% (36)
- 2.3 Cellular Components 2.3.1 Membrane, Cell Wall, and Cytoplasm Davson-Danielli Model of The MembraneDocument16 pages2.3 Cellular Components 2.3.1 Membrane, Cell Wall, and Cytoplasm Davson-Danielli Model of The Membranedavid ngPas encore d'évaluation
- Containing The Cell: Cell Membrane: Part I: Locating Physiology On The Web of KnowledgeDocument5 pagesContaining The Cell: Cell Membrane: Part I: Locating Physiology On The Web of KnowledgeEly TrajicoPas encore d'évaluation
- General Functions and Characteristics: Cell MembraneDocument2 pagesGeneral Functions and Characteristics: Cell MembraneHannaPas encore d'évaluation
- Mccance: Pathophysiology, 6Th Edition: Chapter 01: Cellular Biology Key Points - Print Summary ReviewDocument5 pagesMccance: Pathophysiology, 6Th Edition: Chapter 01: Cellular Biology Key Points - Print Summary ReviewAbiah IsraelPas encore d'évaluation
- 3 Cytology Membran JunctionDocument48 pages3 Cytology Membran JunctionSamPas encore d'évaluation
- Question 1: Identify On A Cell Model or Diagram The Three Major Cell Regions (NucleusDocument10 pagesQuestion 1: Identify On A Cell Model or Diagram The Three Major Cell Regions (NucleusGracea Mae AranetaPas encore d'évaluation
- Cell Membrane (Plasma Membrane)Document6 pagesCell Membrane (Plasma Membrane)Rica NorcioPas encore d'évaluation
- Eukaryotic Cell ComponentsDocument31 pagesEukaryotic Cell ComponentscayetanaramosabejaroPas encore d'évaluation
- Course 2Document28 pagesCourse 2managerw1sbusinessPas encore d'évaluation
- 2 The Cell - Structure and Function 228964Document17 pages2 The Cell - Structure and Function 228964Musharaf RehmanPas encore d'évaluation
- Unit 3: How Are Eukaryotic Cells Organized Into Smaller Parts?Document6 pagesUnit 3: How Are Eukaryotic Cells Organized Into Smaller Parts?juan mondaPas encore d'évaluation
- Histology Text, Problems, TestsDocument334 pagesHistology Text, Problems, TestsmoahedmaedePas encore d'évaluation
- 2 - kh201295 kh201295Document11 pages2 - kh201295 kh201295murtada gubaPas encore d'évaluation
- Cell BiologyDocument3 pagesCell BiologyjamjamjamPas encore d'évaluation
- Basic Cell Theory - StructureDocument32 pagesBasic Cell Theory - StructureRuby SamarPas encore d'évaluation
- Plasma Membrane DeutschDocument5 pagesPlasma Membrane DeutschSafeer SefiPas encore d'évaluation
- Unit 2: Cell Structure and Function Chapters 6-7Document4 pagesUnit 2: Cell Structure and Function Chapters 6-7api-542216138Pas encore d'évaluation
- Human Anatomy 4th Edition Saladin Solutions ManualDocument25 pagesHuman Anatomy 4th Edition Saladin Solutions ManualLeslieBuchanannjfy100% (52)
- CellDocument8 pagesCellvjaPas encore d'évaluation
- BIO 315. NotesDocument42 pagesBIO 315. NotesKevin KipropPas encore d'évaluation
- From Tubulin to Thought: The Nexus of Cytoskeleton Microtubules and Brain Complexity.D'EverandFrom Tubulin to Thought: The Nexus of Cytoskeleton Microtubules and Brain Complexity.Pas encore d'évaluation
- Cell Structure and FunctionDocument9 pagesCell Structure and Functionrbyq9Pas encore d'évaluation
- Prokaryotic CellDocument16 pagesProkaryotic CellKhurram HussainPas encore d'évaluation
- Raven Biology of Plants: Eighth EditionDocument63 pagesRaven Biology of Plants: Eighth EditionMoath EnnabPas encore d'évaluation
- 1.4. Cell StructureDocument4 pages1.4. Cell StructuresamPas encore d'évaluation
- Viii Competitive Cell 1Document25 pagesViii Competitive Cell 1rajesh duaPas encore d'évaluation
- Cell - The Unit of Life - NCERT Based PYQsDocument9 pagesCell - The Unit of Life - NCERT Based PYQsAkhil singhPas encore d'évaluation
- Onion and Human Cheek CellDocument3 pagesOnion and Human Cheek CellAnjehyn Elle100% (1)
- Lab: Onion CellsDocument2 pagesLab: Onion Cellstanatswa100% (1)
- Sperm Morphology - 2019 FinalDocument44 pagesSperm Morphology - 2019 FinalDr Jitu moni DasPas encore d'évaluation
- Introduction To Cell BiologyDocument32 pagesIntroduction To Cell BiologyMustafa Khandgawi100% (3)
- Science Class 7 Week 2 Worksheet 3 PDFDocument2 pagesScience Class 7 Week 2 Worksheet 3 PDFhamza100% (4)
- CheekDocument3 pagesCheekTSha Meunier100% (2)
- LKM 5 - VAKUOLA - Kel 9Document4 pagesLKM 5 - VAKUOLA - Kel 9Farkhatul AzizahPas encore d'évaluation
- Cytoskeleton 2Document7 pagesCytoskeleton 2Basil Francis AlajidPas encore d'évaluation
- 8 Biotechnology ReviewerDocument3 pages8 Biotechnology ReviewerEunice Aruta100% (1)
- Cell DivisionDocument74 pagesCell DivisionNor HilmiPas encore d'évaluation
- Pearson Science TextbookDocument50 pagesPearson Science TextbookRick100% (2)
- 3.4 Structure and Function of Cells JONESDocument5 pages3.4 Structure and Function of Cells JONESJonathan GordonPas encore d'évaluation
- 1.mind Map - Cell - A Unit of LifeDocument100 pages1.mind Map - Cell - A Unit of LifeAyush KumarPas encore d'évaluation
- Grade 7 Unicellular OrganismsDocument10 pagesGrade 7 Unicellular OrganismsGadgetGlitchKillPas encore d'évaluation
- Test Bank For Illustrated Dental Embryology Histology and Anatomy 4th EditionDocument9 pagesTest Bank For Illustrated Dental Embryology Histology and Anatomy 4th Editionpennypowersxbryqnsgtz100% (26)
- Kami Export - Cell Structure Practice (1) - 3Document3 pagesKami Export - Cell Structure Practice (1) - 3Achionta NandyPas encore d'évaluation
- International GCSE Biology Scheme of Work Review 1Document6 pagesInternational GCSE Biology Scheme of Work Review 1Novanolo Christovori ZebuaPas encore d'évaluation
- A1. Illustration of Cell CycleDocument5 pagesA1. Illustration of Cell CycleAllesa San JosePas encore d'évaluation
- Plant Cell Color Page Worksheet and Quiz CeDocument6 pagesPlant Cell Color Page Worksheet and Quiz CeAngel Favila100% (1)
- Session 2 - Cell Structures and Their FunctionsDocument3 pagesSession 2 - Cell Structures and Their FunctionsKrisha Mae PascuaPas encore d'évaluation
- Downloadable Test Bank For Fundamentals of Human Physiology 4th Edition Sherwood FPHP4e - 002TB 1Document22 pagesDownloadable Test Bank For Fundamentals of Human Physiology 4th Edition Sherwood FPHP4e - 002TB 1Zaid Khaled100% (1)
- Chromosome Number Worksheet Meiosis MitosisDocument3 pagesChromosome Number Worksheet Meiosis MitosisDaisy GamerPas encore d'évaluation
- Forest Botany For Forestry StudentsDocument39 pagesForest Botany For Forestry StudentsMadan ThapaPas encore d'évaluation
- Cellular Movement and Muscles: Powerpoint Lecture Slides Prepared by Stephen Gehnrich, Salisbury UniversityDocument89 pagesCellular Movement and Muscles: Powerpoint Lecture Slides Prepared by Stephen Gehnrich, Salisbury UniversityJennie LaoPas encore d'évaluation