Vous aimerez peut-être aussi
- Efectos Del EtilenoDocument4 pagesEfectos Del EtilenolunaPas encore d'évaluation
- El Cuervo - Edgar Allan PoeDocument47 pagesEl Cuervo - Edgar Allan PoeRuth Jhael Mendez ReyesPas encore d'évaluation
- Complicaciones Mediatas de La ExodonciaDocument7 pagesComplicaciones Mediatas de La Exodonciaieshua4572Pas encore d'évaluation
- Torres de AdsorcionDocument7 pagesTorres de AdsorcionCatalina Rojas Correa100% (1)
- NutriciónSaludYalimentosfuncionales DecryptedDocument665 pagesNutriciónSaludYalimentosfuncionales DecryptedCatalina Rojas Correa100% (1)
- Razas Ovinas y Su Rol en Los Sistemas de Cruzamiento OrientaDocument17 pagesRazas Ovinas y Su Rol en Los Sistemas de Cruzamiento OrientaJair Prado RamirezPas encore d'évaluation
- Labores PreparatoriasDocument66 pagesLabores PreparatoriasjotakartikellanPas encore d'évaluation
- Métodos de EsterilizaciónDocument6 pagesMétodos de EsterilizaciónEdith Lopez Gomez100% (1)
- Huesos del Cráneo: Neurocráneo y ViscerocráneoDocument18 pagesHuesos del Cráneo: Neurocráneo y ViscerocráneoFernando Diaz Herrera100% (1)
- Fitohormonas PDFDocument9 pagesFitohormonas PDFJorge RiveraPas encore d'évaluation
- Fisiologia Del Etileno (Unidad IV)Document61 pagesFisiologia Del Etileno (Unidad IV)manuel martinezPas encore d'évaluation
- Antecedentes Del InvernaderoDocument4 pagesAntecedentes Del InvernaderoLizbeth MoralesPas encore d'évaluation
- Etileno regulador claveDocument16 pagesEtileno regulador claveAndrea Mera PalaciosPas encore d'évaluation
- Sesion 5 - EndodonciaDocument32 pagesSesion 5 - EndodonciaValeria Villanueva MontealegrePas encore d'évaluation
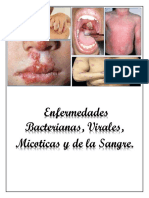
- Enfermedades Virales, Bacterianas y MicoticasDocument36 pagesEnfermedades Virales, Bacterianas y MicoticasJoselin Alexandra Campos MuñozPas encore d'évaluation
- Funciones del etileno en plantasDocument42 pagesFunciones del etileno en plantaslebiatan89100% (1)
- EtilenoDocument46 pagesEtilenoCamilo Andres Benavides MuñozPas encore d'évaluation
- EtilenoDocument13 pagesEtilenomieleritoPas encore d'évaluation
- Causas de La SalinidadDocument27 pagesCausas de La SalinidadCarol ladinoPas encore d'évaluation
- Clasificación de La CariesDocument9 pagesClasificación de La CariesMaria Camila GalloPas encore d'évaluation
- Diagnóstico de Juventud FinalDocument97 pagesDiagnóstico de Juventud FinalchicaquichaPas encore d'évaluation
- Movimientos MandibularesDocument5 pagesMovimientos MandibularesDianela EscalantePas encore d'évaluation
- Componentes y usos de prótesis parcial removibleDocument1 pageComponentes y usos de prótesis parcial removibleOsoMcqueenPas encore d'évaluation
- Disruptores Endocrinos ArticuloDocument6 pagesDisruptores Endocrinos ArticuloOrbis rubraPas encore d'évaluation
- Introducción a la fermentación y ácidos orgánicos producidosDocument27 pagesIntroducción a la fermentación y ácidos orgánicos producidosDerly Loaiza Ninantay100% (1)
- Normas de Bioseguridad InformeDocument4 pagesNormas de Bioseguridad Informemaried eliana marin50% (2)
- Informe - ProteinasDocument8 pagesInforme - ProteinasCintia Azucena Villanueva EstacioPas encore d'évaluation
- Clases de InvernaderosDocument10 pagesClases de InvernaderosCarolGarciaPas encore d'évaluation
- Evaluación de Tierras - ROSSITER PDFDocument49 pagesEvaluación de Tierras - ROSSITER PDFJorge Hernan LondoñoPas encore d'évaluation
- Periodonto NormalDocument2 pagesPeriodonto NormalJeannette BorreroPas encore d'évaluation
- Fotografias Intraorales Forntal y LateralDocument4 pagesFotografias Intraorales Forntal y LateralAnel MamaniPas encore d'évaluation
- Exodoncia en pacientes hipertensosDocument26 pagesExodoncia en pacientes hipertensosWendy Reyes GasteloPas encore d'évaluation
- Topografía en La Agricultura (Autoguardado)Document10 pagesTopografía en La Agricultura (Autoguardado)NoHeliaPeRezPas encore d'évaluation
- Metabolismo Microbiano PDFDocument8 pagesMetabolismo Microbiano PDFHannie BertalotPas encore d'évaluation
- Cancer y Precancer de La Cavidad BucalDocument38 pagesCancer y Precancer de La Cavidad Bucalana Lucia MoscosoPas encore d'évaluation
- Ionomero de VidrioDocument11 pagesIonomero de VidrioAlonso LimachePas encore d'évaluation
- Introduccion Ensayo 10Document7 pagesIntroduccion Ensayo 10santiagoPas encore d'évaluation
- INTRODUCCION TOPOGRAFIA AgronomiaDocument52 pagesINTRODUCCION TOPOGRAFIA AgronomiaClinton MHPas encore d'évaluation
- INFORME 11, Semana 12, Palomino JassirDocument6 pagesINFORME 11, Semana 12, Palomino JassirJassir Palomino SanchezPas encore d'évaluation
- Ciclo de Krebs: Balance energético y regulaciónDocument3 pagesCiclo de Krebs: Balance energético y regulaciónstefanyPas encore d'évaluation
- Urgencia en EndodonciaDocument7 pagesUrgencia en EndodonciaBlas HernandezPas encore d'évaluation
- Enfermedades Relacionadas Con El Metabolismo de Purinas y PirimidinasDocument23 pagesEnfermedades Relacionadas Con El Metabolismo de Purinas y PirimidinasDiana Cárdenas100% (4)
- Exposiicon de EspigosDocument5 pagesExposiicon de EspigosCsAr Zuñiga Lujan100% (2)
- Cuadro Sinóptico - HongosDocument1 pageCuadro Sinóptico - HongosSalito VillacísPas encore d'évaluation
- Aislamiento AbsolutoDocument8 pagesAislamiento AbsolutoJesús LugoPas encore d'évaluation
- Intoxicación Por FluorDocument34 pagesIntoxicación Por FluorKarliilu ZaldiivarPas encore d'évaluation
- Materiales de Impresion DentalDocument3 pagesMateriales de Impresion DentalDulis OrtizPas encore d'évaluation
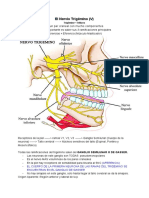
- Par Craneal 5 PDFDocument8 pagesPar Craneal 5 PDFAlicia Trimmiño JaramilloPas encore d'évaluation
- FofatasasDocument17 pagesFofatasasLuis E. Cubas GuevaraPas encore d'évaluation
- MANEJO TERAPEÚTICO DEL PACIENTE CON XEROSTOMÍA PedroDocument16 pagesMANEJO TERAPEÚTICO DEL PACIENTE CON XEROSTOMÍA PedroPedro Sacaico PalaciosPas encore d'évaluation
- Tratamientos de Superficies de ImplantesDocument22 pagesTratamientos de Superficies de ImplantesMelissa Yanira Quiquia CajahuancaPas encore d'évaluation
- Procesamiento de La Película para Radiografía DentalDocument49 pagesProcesamiento de La Película para Radiografía DentalNatalia Mesa de la CruzPas encore d'évaluation
- Resumen Clase 1 - Adhesive MeetingDocument6 pagesResumen Clase 1 - Adhesive MeetingJorge Luis Perez GarcíaPas encore d'évaluation
- Protozoos 110226185943 Phpapp01Document83 pagesProtozoos 110226185943 Phpapp01edwin920710Pas encore d'évaluation
- Cultivo in vitro de tejidos vegetales: historia, tipos y aplicacionesDocument41 pagesCultivo in vitro de tejidos vegetales: historia, tipos y aplicacionesNeco Chan100% (1)
- Filtro de MembranasDocument19 pagesFiltro de MembranasNicole Orellana PintoPas encore d'évaluation
- Proteinas Del HuevoDocument2 pagesProteinas Del HuevoJuan ChingaliPas encore d'évaluation
- Biofilms Dentales. Objetivos Terapéuticos DifícilesDocument44 pagesBiofilms Dentales. Objetivos Terapéuticos DifícilesMacarena Ivonne González Vargas100% (3)
- Globulos RojosDocument15 pagesGlobulos RojosLeonel Fernendez Ramirez100% (1)
- ConductometríaDocument14 pagesConductometríarct85100% (1)
- Historia y características de la gingivitis ulceronecrotizante aguda (GUNADocument45 pagesHistoria y características de la gingivitis ulceronecrotizante aguda (GUNAMoriano MiguelPas encore d'évaluation
- Práctica Nº7 EMULSIONESDocument7 pagesPráctica Nº7 EMULSIONESjudit delgado perezPas encore d'évaluation
- 04 EtilenoDocument9 pages04 EtilenodokygodPas encore d'évaluation
- Cuestionario de PreguntasDocument8 pagesCuestionario de PreguntasLenin Alexander Vargas Bustamante50% (2)
- Informe 4 - Respiración y EtilenoDocument9 pagesInforme 4 - Respiración y EtilenoJoshua Sebastian Vargas NacarinoPas encore d'évaluation
- Curso: Buenas Prácticas de Manufactura (BPM) para La Industria Farmacéutica, Cosmética y Normas HACCP para AlimentosDocument6 pagesCurso: Buenas Prácticas de Manufactura (BPM) para La Industria Farmacéutica, Cosmética y Normas HACCP para AlimentosCatalina Rojas CorreaPas encore d'évaluation
- Guia PhvaDocument11 pagesGuia Phvamiryam timisay vanegas hernandezPas encore d'évaluation
- Sesión 2. Marzo 16 Boydorr PDFDocument27 pagesSesión 2. Marzo 16 Boydorr PDFCatalina Rojas CorreaPas encore d'évaluation
- Formulario - Inscripcion - Dafp PLANTA TEMPORAL Hoja 2 PDFDocument1 pageFormulario - Inscripcion - Dafp PLANTA TEMPORAL Hoja 2 PDFCatalina Rojas CorreaPas encore d'évaluation
- OscarDocument3 pagesOscarCatalina Rojas CorreaPas encore d'évaluation
- Inscribirme PDFDocument1 pageInscribirme PDFCatalina Rojas CorreaPas encore d'évaluation
- Pregradoyposgrado PDFDocument1 pagePregradoyposgrado PDFYeferson GuarinPas encore d'évaluation
- Tarea Poema y FigurasDocument2 pagesTarea Poema y FigurasCatalina Rojas CorreaPas encore d'évaluation
- Tener en CuentaDocument1 pageTener en CuentaCatalina Rojas CorreaPas encore d'évaluation
- ColcienciasDocument10 pagesColcienciasCatalina Rojas CorreaPas encore d'évaluation
- AnálisisDocument1 pageAnálisisCatalina Rojas CorreaPas encore d'évaluation
- Tema 5 PSIA - Isotermas de LagmuirDocument14 pagesTema 5 PSIA - Isotermas de LagmuirJemmy VillalobosPas encore d'évaluation
- M 3874Document1 pageM 3874Catalina Rojas CorreaPas encore d'évaluation
- AnálisisDocument1 pageAnálisisCatalina Rojas CorreaPas encore d'évaluation
- Diseño Metodológico Del Tratamiento de Aguas Residuales en Empresas de Alimentos CárnicosDocument2 pagesDiseño Metodológico Del Tratamiento de Aguas Residuales en Empresas de Alimentos CárnicosCatalina Rojas CorreaPas encore d'évaluation
- Foro Semana 1Document2 pagesForo Semana 1Catalina Rojas CorreaPas encore d'évaluation
- Foro Semana 1Document1 pageForo Semana 1Catalina Rojas CorreaPas encore d'évaluation
- NTC4458Document18 pagesNTC4458Catalina Rojas CorreaPas encore d'évaluation
- 15-Crema para FrutasDocument1 page15-Crema para FrutasCatalina Rojas CorreaPas encore d'évaluation
- NTC4092 Requisitos y Directrices de Laboratorio PDFDocument85 pagesNTC4092 Requisitos y Directrices de Laboratorio PDFJose StewartPas encore d'évaluation
- Actividad Unidad UnoDocument4 pagesActividad Unidad UnoCatalina Rojas CorreaPas encore d'évaluation
- Secuencia de Trabajo Diario Iep Davis Moody Kids Del 20 Al 24 de MarzoDocument4 pagesSecuencia de Trabajo Diario Iep Davis Moody Kids Del 20 Al 24 de MarzoRaquel Nataly Ramos GuerraPas encore d'évaluation
- Arcos Dentarios: Características y FormasDocument57 pagesArcos Dentarios: Características y FormasLeidy UrquidiPas encore d'évaluation
- Parto en CerdasDocument17 pagesParto en CerdasOscar Leonel Torres CruzPas encore d'évaluation
- 2ºficha de Lectura Comprensiva 10Document8 pages2ºficha de Lectura Comprensiva 10Laura FernándezPas encore d'évaluation
- Problemas de Genética 1 PDFDocument2 pagesProblemas de Genética 1 PDFCarlos Ferrer GarcíaPas encore d'évaluation
- Phylum NematomorphaDocument14 pagesPhylum Nematomorphacarolina pradosPas encore d'évaluation
- 3 Orden Squamata PDFDocument42 pages3 Orden Squamata PDFAnonymous BrO7zvF3PxPas encore d'évaluation
- Martorelli, Marcotegui & Alda, 2006Document16 pagesMartorelli, Marcotegui & Alda, 2006Sergio MartorelliPas encore d'évaluation
- GLOSARIO de AnimalesDocument5 pagesGLOSARIO de AnimalesYorela Torres Castillo100% (1)
- Morfologia de InsectosDocument213 pagesMorfologia de InsectosRocio RodriguezPas encore d'évaluation
- Cultivo Del Frijol (Martínez-Unesr)Document9 pagesCultivo Del Frijol (Martínez-Unesr)Fredrick AlvaradoPas encore d'évaluation
- Ciclo Verano 2009 Colegios Bryce – Joyce – Freud ÁlgebraDocument17 pagesCiclo Verano 2009 Colegios Bryce – Joyce – Freud ÁlgebraAurelio BaldorPas encore d'évaluation
- EsmalteDocument2 pagesEsmalteVictor VargasPas encore d'évaluation
- Interacción Entre Las Células y Ambiente II 2016-02Document35 pagesInteracción Entre Las Células y Ambiente II 2016-02jcmv31Pas encore d'évaluation
- Cuento El Rey La RanaDocument8 pagesCuento El Rey La RanaNorma Elizabeth Lopez VillatoroPas encore d'évaluation
- Nutricion Animal ProteínasDocument28 pagesNutricion Animal Proteínasメサ ニフラ デニルソン80% (5)
- El rey de los buitresDocument10 pagesEl rey de los buitresTemploTeolaEscuelaDeMagiaEsotericaPas encore d'évaluation
- OrnitologiaDocument3 pagesOrnitologiaMaritza Moncayo BermudezPas encore d'évaluation
- Bioquímica Centrifugación de Organelos de EspinacaDocument8 pagesBioquímica Centrifugación de Organelos de Espinacaxomi2530Pas encore d'évaluation
- Temas 11 A 15Document19 pagesTemas 11 A 15Paula Frez Pavez100% (1)
- 14 Congreso Etologia. Resumenes 2 PDFDocument95 pages14 Congreso Etologia. Resumenes 2 PDFgonzaloabreaPas encore d'évaluation
- getReporteCalas 14-12-22 EsperanzaDocument2 pagesgetReporteCalas 14-12-22 EsperanzaJosePas encore d'évaluation
- Guia OfidismoDocument78 pagesGuia OfidismoDamiley GHPas encore d'évaluation
- La VacaDocument3 pagesLa Vacawsmr1234Pas encore d'évaluation
- Pie (Articulaciones J Huesos J Musculos) JGSSDocument16 pagesPie (Articulaciones J Huesos J Musculos) JGSSJazz Solis SolanoPas encore d'évaluation
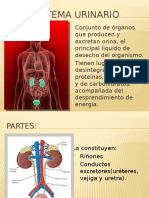
- Sistema UtinarioDocument13 pagesSistema UtinarioYESSICALISBETHTAPas encore d'évaluation
- Taller de Refuerzo I Periodo Biologia SextoDocument4 pagesTaller de Refuerzo I Periodo Biologia SextoRubi Cardenas100% (1)