Vous aimerez peut-être aussi
- A Cross Sectional Study On Bacteriological Profile and Antibiogram of Cholecystitis and Cholelithiasis - IJMM 2021Document7 pagesA Cross Sectional Study On Bacteriological Profile and Antibiogram of Cholecystitis and Cholelithiasis - IJMM 2021International Network For Natural SciencesPas encore d'évaluation
- Structure of Zooplanktonic Communities in Lakes Ossa and Mwembe at Dizangue (Cameroun, Central Africa) - JBES 2021Document15 pagesStructure of Zooplanktonic Communities in Lakes Ossa and Mwembe at Dizangue (Cameroun, Central Africa) - JBES 2021International Network For Natural SciencesPas encore d'évaluation
- Assessment of Bud-Grafting and Side-Grafting Techniques To Support The Improvement of Productivity and Quality of Somatic Embryogenesis Cocoa (Theobroma Cacao L.) in South SulawesiDocument9 pagesAssessment of Bud-Grafting and Side-Grafting Techniques To Support The Improvement of Productivity and Quality of Somatic Embryogenesis Cocoa (Theobroma Cacao L.) in South SulawesiInternational Network For Natural SciencesPas encore d'évaluation
- Growth and Yield Performance of Pechay (Brassica Napus L.) in Hydroponics System As Influenced by Magnetic Field - IJAAR 2021Document8 pagesGrowth and Yield Performance of Pechay (Brassica Napus L.) in Hydroponics System As Influenced by Magnetic Field - IJAAR 2021International Network For Natural SciencesPas encore d'évaluation
- Reuse of Wastewater For Irrigation Purposes - JBES 2021 by INNSPUBDocument12 pagesReuse of Wastewater For Irrigation Purposes - JBES 2021 by INNSPUBInternational Network For Natural SciencesPas encore d'évaluation
- Reuse of Wastewater For Irrigation Purposes - JBES 2021 by INNSPUBDocument12 pagesReuse of Wastewater For Irrigation Purposes - JBES 2021 by INNSPUBInternational Network For Natural SciencesPas encore d'évaluation
- Musa X Paradisiaca Manihot Esculenta FoutouDocument9 pagesMusa X Paradisiaca Manihot Esculenta FoutouInternational Network For Natural SciencesPas encore d'évaluation
- Impact of Phragmanthera Capitata (Sprenge.) Balle On Pod and Beans Production of Two Cocoa Clones in Nkoemvone Seed Fields (South Cameroun) - JBES @scribdDocument9 pagesImpact of Phragmanthera Capitata (Sprenge.) Balle On Pod and Beans Production of Two Cocoa Clones in Nkoemvone Seed Fields (South Cameroun) - JBES @scribdInternational Network For Natural SciencesPas encore d'évaluation
- Vegetative Propagation of Anonidium Mannii (Oliver) Engler & Diels (Annonaceae) by Leafy Stem Cuttings in Kisangani, DR Congo-IJAARDocument11 pagesVegetative Propagation of Anonidium Mannii (Oliver) Engler & Diels (Annonaceae) by Leafy Stem Cuttings in Kisangani, DR Congo-IJAARInternational Network For Natural SciencesPas encore d'évaluation
- Study of Virulence Genes in Vancomycin Resistant Enterococci (Vre) From Animals and Human Clinical Isolates-IJB 2021Document15 pagesStudy of Virulence Genes in Vancomycin Resistant Enterococci (Vre) From Animals and Human Clinical Isolates-IJB 2021International Network For Natural SciencesPas encore d'évaluation
- Call For Paper - IJAAR JournalsDocument1 pageCall For Paper - IJAAR JournalsInternational Network For Natural SciencesPas encore d'évaluation
- Call For Paper 2022 - IJMM Journals by INNSpubDocument1 pageCall For Paper 2022 - IJMM Journals by INNSpubInternational Network For Natural SciencesPas encore d'évaluation
- Ormosia Calavensis Litsea PhilippinensisDocument9 pagesOrmosia Calavensis Litsea PhilippinensisInternational Network For Natural SciencesPas encore d'évaluation
- Meat Tenderization Through Plant Proteases - A Mini Review-IjbDocument13 pagesMeat Tenderization Through Plant Proteases - A Mini Review-IjbInternational Network For Natural SciencesPas encore d'évaluation
- Archive - April Issue 2021 by IJB Journals - INNSPUBDocument4 pagesArchive - April Issue 2021 by IJB Journals - INNSPUBInternational Network For Natural SciencesPas encore d'évaluation
- The Effectiveness of Experience and Nature-Based Learning Activities in Enhancing Students Environmental Attitude - JBES @scribdDocument14 pagesThe Effectiveness of Experience and Nature-Based Learning Activities in Enhancing Students Environmental Attitude - JBES @scribdInternational Network For Natural SciencesPas encore d'évaluation
- Review On: Impact of Seed Rates and Method of Sowing On Yield and Yield Related Traits of Teff (Eragrostis Teff (Zucc.) Trotter) - IJAAR @scribdDocument8 pagesReview On: Impact of Seed Rates and Method of Sowing On Yield and Yield Related Traits of Teff (Eragrostis Teff (Zucc.) Trotter) - IJAAR @scribdInternational Network For Natural SciencesPas encore d'évaluation
- Research Paper: Journal NameDocument10 pagesResearch Paper: Journal NameInternational Network For Natural SciencesPas encore d'évaluation
- Occurrence Level of Urban and Peri-Urban Farming Activities Across Major Network Links of Nairobi County, Kenya - IJAAR @scribdDocument13 pagesOccurrence Level of Urban and Peri-Urban Farming Activities Across Major Network Links of Nairobi County, Kenya - IJAAR @scribdInternational Network For Natural SciencesPas encore d'évaluation
- Performance Assessment of Water Filtration Plants in Pakistan MDocument15 pagesPerformance Assessment of Water Filtration Plants in Pakistan MInternational Network For Natural SciencesPas encore d'évaluation
- Medicinal Plants Used Against Epilepsy by The Local Communities of Sargodha Region, Pakistan-JBESDocument13 pagesMedicinal Plants Used Against Epilepsy by The Local Communities of Sargodha Region, Pakistan-JBESInternational Network For Natural SciencesPas encore d'évaluation
- Carbonized Rice Hull (CRH) Silica Extract Efficiency For Yield and Nutrient Uptake of Cucumber - IJB JournalDocument7 pagesCarbonized Rice Hull (CRH) Silica Extract Efficiency For Yield and Nutrient Uptake of Cucumber - IJB JournalInternational Network For Natural SciencesPas encore d'évaluation
- The Effect of Seed Coat Removal On Seed Germination of Terminalia Superba Engl. & Diels-JBESDocument10 pagesThe Effect of Seed Coat Removal On Seed Germination of Terminalia Superba Engl. & Diels-JBESInternational Network For Natural Sciences100% (1)
- Impacts of Management and Modernization On Water Savings in Large Irrigation SystemsDocument11 pagesImpacts of Management and Modernization On Water Savings in Large Irrigation SystemsInternational Network For Natural SciencesPas encore d'évaluation
- Identification and Pharmacological Properties of Coastal Plants From Barangay Diguisit, Baler, Aurora - JBES-Vol-18-No-6Document11 pagesIdentification and Pharmacological Properties of Coastal Plants From Barangay Diguisit, Baler, Aurora - JBES-Vol-18-No-6International Network For Natural SciencesPas encore d'évaluation
- Productivity and Quality of Sugar Beet As Affecting by Sowing and Irrigation Methods and Hill Spacings-IJAARDocument11 pagesProductivity and Quality of Sugar Beet As Affecting by Sowing and Irrigation Methods and Hill Spacings-IJAARInternational Network For Natural SciencesPas encore d'évaluation
- Findings From A Survey in Western Kenya To Determine The Soil Fertility Replenishment Technologies-IJAAR - INNSPUBDocument11 pagesFindings From A Survey in Western Kenya To Determine The Soil Fertility Replenishment Technologies-IJAAR - INNSPUBInternational Network For Natural SciencesPas encore d'évaluation
- Effectiveness of Community Forest Association and Water Resource Users' Associations in Discharging Their Statutory Functions: A Case of Kilungu Catchment, Kenya - JBES - INNSPUBDocument16 pagesEffectiveness of Community Forest Association and Water Resource Users' Associations in Discharging Their Statutory Functions: A Case of Kilungu Catchment, Kenya - JBES - INNSPUBInternational Network For Natural SciencesPas encore d'évaluation
- Study of Virulence Genes in Vancomycin Resistant Enterococci (Vre) From Animals and Human Clinical - IJBDocument16 pagesStudy of Virulence Genes in Vancomycin Resistant Enterococci (Vre) From Animals and Human Clinical - IJBInternational Network For Natural SciencesPas encore d'évaluation
- Prediction and Evaluation of Deleterious Non-Synonymous SNPs (NSSNPS) in Human IL17A Gene - IJB-2021Document12 pagesPrediction and Evaluation of Deleterious Non-Synonymous SNPs (NSSNPS) in Human IL17A Gene - IJB-2021International Network For Natural SciencesPas encore d'évaluation
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5794)
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (895)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (400)
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (588)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (74)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (344)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (121)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- KL Trial SPM 2012 Biology k123 SkemaDocument21 pagesKL Trial SPM 2012 Biology k123 SkemasawPas encore d'évaluation
- Week 2 Axial Skeleton (A) Lecture 2Document35 pagesWeek 2 Axial Skeleton (A) Lecture 2Fathimath iba AhmedPas encore d'évaluation
- Natural Sciences 4Document22 pagesNatural Sciences 4Ramin Zand Isa Khani100% (2)
- Gr5 Ibt Eng PracticeDocument7 pagesGr5 Ibt Eng PracticeArmaan50% (2)
- Shark Key LabDocument4 pagesShark Key LabcrnoblePas encore d'évaluation
- Animal Thematic 1852 - 2011: Tony BrayDocument227 pagesAnimal Thematic 1852 - 2011: Tony Braydeepheat_008Pas encore d'évaluation
- Parental Care in Fishes (With Diagram) - Vertebrates - Chordata - ZoologyDocument12 pagesParental Care in Fishes (With Diagram) - Vertebrates - Chordata - ZoologyEr Purushottam Pal100% (1)
- Gill SlitDocument2 pagesGill SlitNirmal BhowmickPas encore d'évaluation
- Adiya 19 AprDocument4 pagesAdiya 19 AprAyu DeviPas encore d'évaluation
- Animal MeiosisDocument32 pagesAnimal MeiosisJeremy YuagaPas encore d'évaluation
- Frogdissectionpre Lab SiaDocument2 pagesFrogdissectionpre Lab Siaapi-347123148Pas encore d'évaluation
- Listening EPN 2 Track 17 N 18 Disc 1 IntermediateDocument2 pagesListening EPN 2 Track 17 N 18 Disc 1 IntermediatetriasyogiPas encore d'évaluation
- A List of OlogiesDocument2 pagesA List of OlogiesVishal HemnaniPas encore d'évaluation
- Bones Forming The JointDocument29 pagesBones Forming The JointvrajPas encore d'évaluation
- Report Text About PandaDocument2 pagesReport Text About PandaYOHANES NONOPas encore d'évaluation
- Cell: The Fundamental Unit of Life: Cell Life Cell Division Multicellular Organisms Egg Energy Flow Metabolism DNADocument8 pagesCell: The Fundamental Unit of Life: Cell Life Cell Division Multicellular Organisms Egg Energy Flow Metabolism DNAQadirPas encore d'évaluation
- Chapter 24. Sense Organs: ObjectivesDocument25 pagesChapter 24. Sense Organs: ObjectivesFrancisco GonzálezPas encore d'évaluation
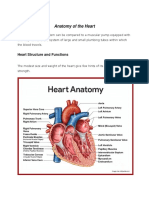
- Anatomy of The HeartDocument18 pagesAnatomy of The Heartvanessaresullar100% (1)
- Chordata NotesDocument5 pagesChordata NotesAbhishek Isaac MathewPas encore d'évaluation
- Walker County Lake Birding ChecklistDocument1 pageWalker County Lake Birding ChecklistHap WalkerPas encore d'évaluation
- Why Are Cervicals Adjust Prone or Seated in GonsteadDocument16 pagesWhy Are Cervicals Adjust Prone or Seated in Gonsteadgonsteadtruth100% (3)
- University of Central Punjab: Paper: Pharmaceutical Biochemistry-IIDocument3 pagesUniversity of Central Punjab: Paper: Pharmaceutical Biochemistry-IIM Imran SajidPas encore d'évaluation
- Soalan Akhir Tahun ENGLISH Kefungsian RendahDocument9 pagesSoalan Akhir Tahun ENGLISH Kefungsian RendahBahnu JanuPas encore d'évaluation
- Cell Transport Practice TestDocument4 pagesCell Transport Practice TestLavander Blush100% (2)
- Comparative and Superlative Adjectives Spi 0501.1.3Document14 pagesComparative and Superlative Adjectives Spi 0501.1.3Aulia Agustin SalimPas encore d'évaluation
- Summary of Germ LayersDocument2 pagesSummary of Germ Layersaichiii.bearPas encore d'évaluation
- Bacterial Anatomy & PhysiologyDocument53 pagesBacterial Anatomy & PhysiologylathaPas encore d'évaluation
- Muscular TissueDocument49 pagesMuscular TissueSteph VeePas encore d'évaluation
- Phylum Cnidaria (Notes)Document1 pagePhylum Cnidaria (Notes)Kristine Claire AlberioPas encore d'évaluation
- The Evolution of Whales: Geology 331Document33 pagesThe Evolution of Whales: Geology 331Emilio Lecaros BustamantePas encore d'évaluation