Vous aimerez peut-être aussi
- Fausto Sterling Gender Sex Sexual Orientation and Identity Are in The Body How Did They Get ThereDocument28 pagesFausto Sterling Gender Sex Sexual Orientation and Identity Are in The Body How Did They Get ThereOntologiaPas encore d'évaluation
- Intersex Concept of Multiple Sexes Is Not NewDocument2 pagesIntersex Concept of Multiple Sexes Is Not NewOntologiaPas encore d'évaluation
- Fausto-Sterling-How Else Can We Study Sex Differences in Early InfancyDocument13 pagesFausto-Sterling-How Else Can We Study Sex Differences in Early InfancyOntologiaPas encore d'évaluation
- Intersex Concept of Multiple Sexes Is Not NewDocument2 pagesIntersex Concept of Multiple Sexes Is Not NewOntologiaPas encore d'évaluation
- Advances in Health Psychology Critical ApproachesDocument3 pagesAdvances in Health Psychology Critical ApproachesOntologiaPas encore d'évaluation
- Biology and IdentityDocument24 pagesBiology and IdentityOntologiaPas encore d'évaluation
- Sexing The Baby Part 1Document10 pagesSexing The Baby Part 1OntologiaPas encore d'évaluation
- Sex Genes A Critical Sociomaterial Approach To The Politics and Molecular Genetics of SexDocument35 pagesSex Genes A Critical Sociomaterial Approach To The Politics and Molecular Genetics of SexOntologiaPas encore d'évaluation
- Neurofeminism and Feminist Neurosciences A Critical Review of Contemporary Brain ResearchDocument10 pagesNeurofeminism and Feminist Neurosciences A Critical Review of Contemporary Brain ResearchOntologiaPas encore d'évaluation
- Central Gender Theoretical Concepts in Health ResearchDocument6 pagesCentral Gender Theoretical Concepts in Health ResearchOntologiaPas encore d'évaluation
- Gender and Discipline in 5-12 Month Old InfantsDocument11 pagesGender and Discipline in 5-12 Month Old InfantsOntologiaPas encore d'évaluation
- Fausto Sterling Frameworks of DesireDocument12 pagesFausto Sterling Frameworks of DesireOntologiaPas encore d'évaluation
- Biocultural Approach The Essence of Anthropological Study in The 21 CenturyDocument12 pagesBiocultural Approach The Essence of Anthropological Study in The 21 CenturyOntologiaPas encore d'évaluation
- Neurofeminism Issues at The Intersection of Feminist TheoryDocument5 pagesNeurofeminism Issues at The Intersection of Feminist TheoryOntologiaPas encore d'évaluation
- Deconstructing Developmental Psychology - Reflections After 20 YearsDocument3 pagesDeconstructing Developmental Psychology - Reflections After 20 YearsOntologiaPas encore d'évaluation
- Nder Psychological PerspectivesDocument4 pagesNder Psychological PerspectivesOntologiaPas encore d'évaluation
- Gender and Culture in Psychology Theories and PracticesDocument4 pagesGender and Culture in Psychology Theories and PracticesOntologiaPas encore d'évaluation
- Gender and Colonialism A Psychological AnalysisDocument5 pagesGender and Colonialism A Psychological AnalysisOntologiaPas encore d'évaluation
- Ainstorm The Flaws in The Science of Sex DifferencesDocument3 pagesAinstorm The Flaws in The Science of Sex DifferencesOntologiaPas encore d'évaluation
- Biosocial Relations of ProductionDocument26 pagesBiosocial Relations of ProductionOntologiaPas encore d'évaluation
- Dynamis Systems Theory Chapter BookDocument55 pagesDynamis Systems Theory Chapter BookManuel Sanchez LopezPas encore d'évaluation
- Rethinking Systems Theory The Problem of CultureDocument24 pagesRethinking Systems Theory The Problem of CultureOntologiaPas encore d'évaluation
- Gender, Nature and CultureDocument58 pagesGender, Nature and CultureOntologiaPas encore d'évaluation
- R.the Oxford Handbook of Feminist Multicultural Counseling PsychologyDocument3 pagesR.the Oxford Handbook of Feminist Multicultural Counseling PsychologyOntologiaPas encore d'évaluation
- Contemporary Feminism For Gender ResearchersDocument5 pagesContemporary Feminism For Gender ResearchersOntologiaPas encore d'évaluation
- R.mediating Race, Globalizing GenderDocument3 pagesR.mediating Race, Globalizing GenderOntologiaPas encore d'évaluation
- Not Your Grandma's Genetics - Some Theoretical NotesDocument9 pagesNot Your Grandma's Genetics - Some Theoretical NotesOntologiaPas encore d'évaluation
- The Five Sexes: Beyond Male and FemaleDocument13 pagesThe Five Sexes: Beyond Male and FemalePNirmalPas encore d'évaluation
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5784)
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (890)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (399)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (265)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (72)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (344)
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (119)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- Klasifikasi NeoplasmaDocument22 pagesKlasifikasi Neoplasmaoscar putraPas encore d'évaluation
- Medical ChemistryDocument32 pagesMedical Chemistryabhishek kumar singhPas encore d'évaluation
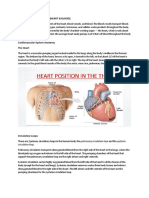
- Cardiovascular SystemDocument17 pagesCardiovascular SystemPoojaPas encore d'évaluation
- Ch. 25 Body Defence MechanismsDocument34 pagesCh. 25 Body Defence Mechanisms吴昊Pas encore d'évaluation
- Erwin-Sheppard Autopsy Report - Redacted by Medical ExaminerDocument17 pagesErwin-Sheppard Autopsy Report - Redacted by Medical ExaminerCheryl Sullenger100% (1)
- Biology: University of Cambridge International Examinations General Certificate of Education Advanced LevelDocument8 pagesBiology: University of Cambridge International Examinations General Certificate of Education Advanced LevelwegdanPas encore d'évaluation
- Motor Speech Disorders - The Handbook of Phonetic Sciences - Blackwell Reference OnlineDocument21 pagesMotor Speech Disorders - The Handbook of Phonetic Sciences - Blackwell Reference OnlineHAKAN ÖZTEMÜRPas encore d'évaluation
- A2 Biology Notes For EdexcelDocument150 pagesA2 Biology Notes For EdexcelAbdulrahman Jijawi83% (6)
- Zincogen NBFDocument7 pagesZincogen NBFcodesolutionroPas encore d'évaluation
- Excretory System of FrogDocument3 pagesExcretory System of FrogSree Lekshmi V SPas encore d'évaluation
- Acceptability: ©WHO/Sergey VolkovDocument26 pagesAcceptability: ©WHO/Sergey VolkovBeatriz PatricioPas encore d'évaluation
- Sonatherapy, Cymatics, and Phonon Chains PDFDocument11 pagesSonatherapy, Cymatics, and Phonon Chains PDFGary Robert Buchanan100% (2)
- Stress 2Document4 pagesStress 2le thi lan anhPas encore d'évaluation
- Sampling and Laboratory Analysis ProceduresDocument22 pagesSampling and Laboratory Analysis ProceduresFauzan HardiPas encore d'évaluation
- Kar Gut Kar 2017Document9 pagesKar Gut Kar 2017Fake TomPas encore d'évaluation
- Introduction BioinformaticsDocument155 pagesIntroduction BioinformaticsMarcus Barros BragaPas encore d'évaluation
- Module 7: Behaviorism Theories of Pavlov, Thorndike, Watson and SkinnerDocument18 pagesModule 7: Behaviorism Theories of Pavlov, Thorndike, Watson and SkinnerAntolin Tambong100% (1)
- 4 Cementum and Alveolar BoneDocument24 pages4 Cementum and Alveolar Boneceudmd3d100% (1)
- Life Processes Class 10 Notes BiologyDocument8 pagesLife Processes Class 10 Notes BiologyEashurock KnigamPas encore d'évaluation
- Cepheids.: No. of Systematic Area Position Variables DeviationDocument9 pagesCepheids.: No. of Systematic Area Position Variables DeviationLefrina GusrianiiPas encore d'évaluation
- Chem 160.1 Post LabDocument7 pagesChem 160.1 Post LabMarlyn AmantePas encore d'évaluation
- Chandini ThesisDocument71 pagesChandini Thesismallesh D trigarPas encore d'évaluation
- Muscular SystemDocument5 pagesMuscular SystemBPas encore d'évaluation
- ANPH 111 (Anatomy and Physiology) : Bachelor of Science in NursingDocument11 pagesANPH 111 (Anatomy and Physiology) : Bachelor of Science in NursingMary Grace MapulaPas encore d'évaluation
- 20-Advances in Cognitive TheoryDocument24 pages20-Advances in Cognitive TheoryAndrea ZeballosPas encore d'évaluation
- Biodegradation of Agricultural Wastes (Rice Straw and Sorghum Stalk) Into Substrates of Utilizable Products Using White Rot FungusDocument7 pagesBiodegradation of Agricultural Wastes (Rice Straw and Sorghum Stalk) Into Substrates of Utilizable Products Using White Rot FungusInes HrastinskiPas encore d'évaluation
- Dwnload Full Medical Terminology For Health Professions 7th Edition Ehrlich Test Bank PDFDocument36 pagesDwnload Full Medical Terminology For Health Professions 7th Edition Ehrlich Test Bank PDFmarchingpalmerbabg100% (9)
- Grade7 LifeScience NGSS 2016 PDFDocument9 pagesGrade7 LifeScience NGSS 2016 PDFJomar SolivaPas encore d'évaluation
- IP DP PSOW - Master ListDocument10 pagesIP DP PSOW - Master ListsilviaPas encore d'évaluation
- Cystic Fibrosis: Nature Reviews Disease Primers May 2015Document20 pagesCystic Fibrosis: Nature Reviews Disease Primers May 2015Tapan AnkleshwariaPas encore d'évaluation