Vous aimerez peut-être aussi
- MCN Drill AnswersDocument12 pagesMCN Drill AnswersHerne Balberde100% (1)
- Your Free Buyer Persona TemplateDocument8 pagesYour Free Buyer Persona Templateel_nakdjoPas encore d'évaluation
- Diode ExercisesDocument5 pagesDiode ExercisesbruhPas encore d'évaluation
- Clearing Negative SpiritsDocument6 pagesClearing Negative SpiritsmehorseblessedPas encore d'évaluation
- Theater 10 Syllabus Printed PDFDocument7 pagesTheater 10 Syllabus Printed PDFJim QuentinPas encore d'évaluation
- Understanding and Targeting Cancer Stem Cells Therapeutic Implications and ChallengesDocument9 pagesUnderstanding and Targeting Cancer Stem Cells Therapeutic Implications and ChallengesMariano PerezPas encore d'évaluation
- Breast Cancer Stem CellsDocument6 pagesBreast Cancer Stem CellsTri Cahya MettaPas encore d'évaluation
- Frontiers in Bioscience E4, 2502-2514, June 1, 2012Document13 pagesFrontiers in Bioscience E4, 2502-2514, June 1, 2012ginocolaciccoPas encore d'évaluation
- Cancer Stem Cells Problems For TherapyDocument15 pagesCancer Stem Cells Problems For Therapyapi-162161895Pas encore d'évaluation
- Cancer Stem Cells: From Birth To DeathDocument30 pagesCancer Stem Cells: From Birth To DeathDito AnurogoPas encore d'évaluation
- Cancer Stem Cells ThesisDocument8 pagesCancer Stem Cells Thesisafknekkfs100% (1)
- Cancer Stem Cell Hypothesis: Implication For Cancer Prevention and TreatmentDocument16 pagesCancer Stem Cell Hypothesis: Implication For Cancer Prevention and TreatmentlaurentiaPas encore d'évaluation
- Stem Cells in Breast Tumours - Are They Ready For The Clinicď Ą, 2012Document13 pagesStem Cells in Breast Tumours - Are They Ready For The Clinicď Ą, 2012Daniela GologanPas encore d'évaluation
- Cancer Stem CellsDocument14 pagesCancer Stem CellsShivani SinghPas encore d'évaluation
- Luna 2016Document34 pagesLuna 2016Jocilene Dantas Torres NascimentoPas encore d'évaluation
- The Tumorigenic CD44Document2 pagesThe Tumorigenic CD44Sulchan Chris WardanaPas encore d'évaluation
- Stem CellsDocument16 pagesStem CellsdregopokePas encore d'évaluation
- The Hypoxic Tumor Microenvironment in Vivo Selects The Cancer Stem Cell Fate of Breast Cancer CellsDocument15 pagesThe Hypoxic Tumor Microenvironment in Vivo Selects The Cancer Stem Cell Fate of Breast Cancer CellsNeha MasarkarPas encore d'évaluation
- Breast Cancer and Cancer Stem Cells: A Mini-Review: Tu Mo Ri, 100: 363-369, 2014Document7 pagesBreast Cancer and Cancer Stem Cells: A Mini-Review: Tu Mo Ri, 100: 363-369, 2014Ava BaranPas encore d'évaluation
- 1 s2.0 S1044579X22002644 Main 1Document15 pages1 s2.0 S1044579X22002644 Main 1ali69rmrimsPas encore d'évaluation
- Monocytes in Tumorigenesis and Tumor ImmunotherapyDocument19 pagesMonocytes in Tumorigenesis and Tumor ImmunotherapyIvan PerezPas encore d'évaluation
- Cancer Stem Cells in Colorectal Cancer and The Association With Chemotherapy ResistanceDocument13 pagesCancer Stem Cells in Colorectal Cancer and The Association With Chemotherapy ResistanceFarmácia Oncológica HC - PEPas encore d'évaluation
- Cancer Stem CellDocument21 pagesCancer Stem CellHakim Asif HaiderPas encore d'évaluation
- Cancer y Celulas Madre 2022Document11 pagesCancer y Celulas Madre 2022Jose EdgarPas encore d'évaluation
- Stem Cell Concepts Renew Cancer Research: ASH 50th Anniversary ReviewDocument15 pagesStem Cell Concepts Renew Cancer Research: ASH 50th Anniversary ReviewbloodbenderPas encore d'évaluation
- Cancers: Circulating Tumor Cells in Prostate CancerDocument15 pagesCancers: Circulating Tumor Cells in Prostate CancerMônicaDíazPas encore d'évaluation
- 1-0articulo MM, PiRNA-823 (Myeloid-Derived Suppressor Cells Endow)Document12 pages1-0articulo MM, PiRNA-823 (Myeloid-Derived Suppressor Cells Endow)Jhonathan David Guevara RamirezPas encore d'évaluation
- Clinics Oncology: Circulating Tumor Cells: One Step Ahead of Precision MedicineDocument3 pagesClinics Oncology: Circulating Tumor Cells: One Step Ahead of Precision MedicineClinics OncologyPas encore d'évaluation
- Qiao 2016Document10 pagesQiao 2016Eliza TotePas encore d'évaluation
- Stem Cells and Colorectal CarcinogenesisDocument6 pagesStem Cells and Colorectal Carcinogenesislavinia diaPas encore d'évaluation
- Celular Dormancy MechanismsDocument17 pagesCelular Dormancy MechanismsJoão FrancoPas encore d'évaluation
- Getting To The Stem of CancerDocument2 pagesGetting To The Stem of CancerEduardo SautchukPas encore d'évaluation
- Stem CellsDocument9 pagesStem CellsMaria Del Mar RoblesPas encore d'évaluation
- MamosphereDocument10 pagesMamospheremuruganvvkPas encore d'évaluation
- Pancreas e Cell StaminaliDocument4 pagesPancreas e Cell StaminaliAntonio MichelottiPas encore d'évaluation
- Cancers 12 03581 v2Document25 pagesCancers 12 03581 v2Danna VeraPas encore d'évaluation
- Hallmarks of Cancer: The Next Generation: ReviewDocument29 pagesHallmarks of Cancer: The Next Generation: Review122307403Pas encore d'évaluation
- MSC e TumorDocument12 pagesMSC e Tumorargolo_napoleaoPas encore d'évaluation
- Cancer Stem Cell Niche The Place To BeDocument7 pagesCancer Stem Cell Niche The Place To BeAlejandra Isabel Juarez JimenezPas encore d'évaluation
- Cancer-Associated Fibroblasts in Tumor Microenvironment - Accomplices in Tumor Malignancy - ScienceDirectDocument6 pagesCancer-Associated Fibroblasts in Tumor Microenvironment - Accomplices in Tumor Malignancy - ScienceDirectPedro MatosPas encore d'évaluation
- Literature Review Breast Cancer PDFDocument7 pagesLiterature Review Breast Cancer PDFaflsimgfs100% (1)
- Human Breast Cancer Stem Cell Markers CD44 and CD24: Enriching For Cells With Functional Properties in Mice or in Man?Document3 pagesHuman Breast Cancer Stem Cell Markers CD44 and CD24: Enriching For Cells With Functional Properties in Mice or in Man?Nehaa RoyPas encore d'évaluation
- Ranking Ativação WNTDocument11 pagesRanking Ativação WNTGabriel LongoPas encore d'évaluation
- BMRI2020-6051210Human Organ-Specific 3D Cancer Models Produced by TheDocument23 pagesBMRI2020-6051210Human Organ-Specific 3D Cancer Models Produced by TheSergio CasadiegosPas encore d'évaluation
- Cas 13155Document7 pagesCas 13155Dana LebadaPas encore d'évaluation
- Cancer Stem CellsDocument9 pagesCancer Stem CellsBasant Hamdy AbuZaidPas encore d'évaluation
- Stem CellDocument13 pagesStem CellVinnie Juliana YonatanPas encore d'évaluation
- Seminars in Cancer Biology: Eleni Mavrogonatou, Harris Pratsinis, Dimitris KletsasDocument10 pagesSeminars in Cancer Biology: Eleni Mavrogonatou, Harris Pratsinis, Dimitris KletsasMIGUEL MORENOPas encore d'évaluation
- 1001Document15 pages1001BinoPas encore d'évaluation
- Pathophysiology and Biology of Peritoneal Carcinomatosis: Antonio Macrì, MD, ProfessorDocument7 pagesPathophysiology and Biology of Peritoneal Carcinomatosis: Antonio Macrì, MD, ProfessorJoia De LeonPas encore d'évaluation
- Ijms 21 04521Document19 pagesIjms 21 04521Farah BdrPas encore d'évaluation
- Identification of Novel Drugs To Target Dormant MicrometastasesDocument12 pagesIdentification of Novel Drugs To Target Dormant MicrometastasesRoman TilahunPas encore d'évaluation
- Fulltext 18Document4 pagesFulltext 18Eli KhPas encore d'évaluation
- Mesenchymal Stem Cell Mediated Cancer Therapy CryovidaDocument8 pagesMesenchymal Stem Cell Mediated Cancer Therapy CryovidaJob MonobePas encore d'évaluation
- Koshkin 2016Document10 pagesKoshkin 2016chemistpl420Pas encore d'évaluation
- Topics: Nasopharyngeal CarcinomaDocument21 pagesTopics: Nasopharyngeal CarcinomaShruthi Y nursingPas encore d'évaluation
- Kim 2015Document8 pagesKim 2015ingePas encore d'évaluation
- FlowcytometryDocument10 pagesFlowcytometryWidia MeliawatiPas encore d'évaluation
- Hallmarks of Cancer: The Next GenerationDocument6 pagesHallmarks of Cancer: The Next GenerationKaren Joy MariñasPas encore d'évaluation
- The Cancer HandbookDocument1 529 pagesThe Cancer HandbookAsimaJulianaSiregarPas encore d'évaluation
- Tmp146e TMPDocument6 pagesTmp146e TMPFrontiersPas encore d'évaluation
- 3-Mesenchimal Stem Cell DevelopmentDocument10 pages3-Mesenchimal Stem Cell Developmentabdulloh suyutiPas encore d'évaluation
- The Identi Fications and Clinical Implications of Cancer Stem Cells in Colorectal CancerDocument10 pagesThe Identi Fications and Clinical Implications of Cancer Stem Cells in Colorectal CancerManuel GuachoPas encore d'évaluation
- Accessories To The Crime Functions of Cells Recruited To The Tumor MicroenvironmentDocument14 pagesAccessories To The Crime Functions of Cells Recruited To The Tumor MicroenvironmentMariano PerezPas encore d'évaluation
- ShinMoe MorningIntroDocument143 pagesShinMoe MorningIntroApplePas encore d'évaluation
- Introduction To Critical Thinking V1Document1 pageIntroduction To Critical Thinking V1ApplePas encore d'évaluation
- Rockwood (2011) - Materials Fabrication From Bombyx Mori Silk Fibroin - Nature ProtocolsDocument20 pagesRockwood (2011) - Materials Fabrication From Bombyx Mori Silk Fibroin - Nature ProtocolsApplePas encore d'évaluation
- Coordination Comp'd ExerciseDocument4 pagesCoordination Comp'd Exercisealvin2282Pas encore d'évaluation
- The Role of The BRCA1 Tumor Suppressor in DNA Double-Strand Break RepairDocument9 pagesThe Role of The BRCA1 Tumor Suppressor in DNA Double-Strand Break RepairApplePas encore d'évaluation
- Concentrating Partial Entanglement by Local OperationsDocument21 pagesConcentrating Partial Entanglement by Local OperationsbhpliaoPas encore d'évaluation
- Focus Group DiscussionDocument13 pagesFocus Group DiscussionSumon ChowdhuryPas encore d'évaluation
- The Recipe For Oleander Sou1Document4 pagesThe Recipe For Oleander Sou1Anthony SullivanPas encore d'évaluation
- Simple Future Tense & Future Continuous TenseDocument2 pagesSimple Future Tense & Future Continuous TenseFarris Ab RashidPas encore d'évaluation
- Yukot,+houkelin 2505 11892735 Final+Paper+Group+41Document17 pagesYukot,+houkelin 2505 11892735 Final+Paper+Group+410191720003 ELIAS ANTONIO BELLO LEON ESTUDIANTE ACTIVOPas encore d'évaluation
- Nastran 2012 Superelements UgDocument974 pagesNastran 2012 Superelements Ugds_srinivasPas encore d'évaluation
- A Passage To AfricaDocument25 pagesA Passage To AfricaJames Reinz100% (2)
- FMEA 4th BOOK PDFDocument151 pagesFMEA 4th BOOK PDFLuis Cárdenas100% (2)
- Romanian Oil IndustryDocument7 pagesRomanian Oil IndustryEnot SoulavierePas encore d'évaluation
- Aspects of The Language - Wintergirls Attached File 3Document17 pagesAspects of The Language - Wintergirls Attached File 3api-207233303Pas encore d'évaluation
- Windows SCADA Disturbance Capture: User's GuideDocument23 pagesWindows SCADA Disturbance Capture: User's GuideANDREA LILIANA BAUTISTA ACEVEDOPas encore d'évaluation
- Lesson 3 The Prodigal SonDocument2 pagesLesson 3 The Prodigal Sonapi-241115908Pas encore d'évaluation
- Work Teams and GroupsDocument6 pagesWork Teams and GroupsFides AvendanPas encore d'évaluation
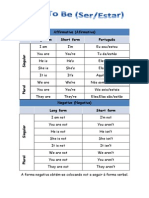
- Affirmative (Afirmativa) Long Form Short Form PortuguêsDocument3 pagesAffirmative (Afirmativa) Long Form Short Form PortuguêsAnitaYangPas encore d'évaluation
- The Changeling by Thomas MiddletonDocument47 pagesThe Changeling by Thomas MiddletonPaulinaOdeth RothPas encore d'évaluation
- Nature, and The Human Spirit: A Collection of QuotationsDocument2 pagesNature, and The Human Spirit: A Collection of QuotationsAxl AlfonsoPas encore d'évaluation
- GearsDocument14 pagesGearsZulhilmi Chik TakPas encore d'évaluation
- Springfield College Lesson Plan Template PHED 237: The Learning and Performance of Physical ActivitiesDocument5 pagesSpringfield College Lesson Plan Template PHED 237: The Learning and Performance of Physical Activitiesapi-285421100Pas encore d'évaluation
- IBM System X UPS Guide v1.4.0Document71 pagesIBM System X UPS Guide v1.4.0Phil JonesPas encore d'évaluation
- Maria MakilingDocument2 pagesMaria MakilingRommel Villaroman Esteves0% (1)
- SassigbmentDocument7 pagesSassigbmentFurkaan Ali KhanPas encore d'évaluation
- Accounting 110: Acc110Document19 pagesAccounting 110: Acc110ahoffm05100% (1)
- ACE Resilience Questionnaires Derek Farrell 2Document6 pagesACE Resilience Questionnaires Derek Farrell 2CATALINA UNDURRAGA UNDURRAGAPas encore d'évaluation
- A Study of Consumer Behavior in Real Estate Sector: Inderpreet SinghDocument17 pagesA Study of Consumer Behavior in Real Estate Sector: Inderpreet SinghMahesh KhadePas encore d'évaluation
- 9m.2-L.5@i Have A Dream & Literary DevicesDocument2 pages9m.2-L.5@i Have A Dream & Literary DevicesMaria BuizonPas encore d'évaluation