Vous aimerez peut-être aussi
- Retinopathy of PrematurityDocument20 pagesRetinopathy of PrematurityChikita Artia SariPas encore d'évaluation
- Retinal DetachmentDocument14 pagesRetinal DetachmentChikita Artia Sari100% (1)
- Hypertensive RetinopathyDocument27 pagesHypertensive RetinopathyChikita Artia SariPas encore d'évaluation
- Optic NeuritisDocument25 pagesOptic NeuritisChikita Artia SariPas encore d'évaluation
- Cover 2Document1 pageCover 2Chikita Artia SariPas encore d'évaluation
- EPISCLERITIS & ScleritisDocument14 pagesEPISCLERITIS & ScleritisChikita Artia SariPas encore d'évaluation
- Case Report Mature Cataract: Created By: Chikita Artia Sari I 11109014Document1 pageCase Report Mature Cataract: Created By: Chikita Artia Sari I 11109014Chikita Artia SariPas encore d'évaluation
- ROP Paeds PresentDocument16 pagesROP Paeds PresentChikita Artia SariPas encore d'évaluation
- Photophobia in Viral Conjunctivitis: Homework Ophtalmology DepartementDocument1 pagePhotophobia in Viral Conjunctivitis: Homework Ophtalmology DepartementChikita Artia SariPas encore d'évaluation
- Metoprolol Succinate Therapy Associated With Erythema MultiformeDocument2 pagesMetoprolol Succinate Therapy Associated With Erythema MultiformeChikita Artia SariPas encore d'évaluation
- Determination of Statement of Principles Concerning PHOTOCONTACT DERMATITISDocument3 pagesDetermination of Statement of Principles Concerning PHOTOCONTACT DERMATITISChikita Artia SariPas encore d'évaluation
- Varicella Zooster VirusDocument9 pagesVaricella Zooster VirusChikita Artia SariPas encore d'évaluation
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (588)
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5795)
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (895)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (345)
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (400)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (74)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (266)
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (121)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- Ashkenazi Jewish ScreeningDocument10 pagesAshkenazi Jewish ScreeningJorge SHPas encore d'évaluation
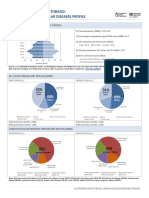
- Trinidad and Tobago: Cardiovascular Diseases ProfileDocument2 pagesTrinidad and Tobago: Cardiovascular Diseases ProfileKay BristolPas encore d'évaluation
- Meditation Its EffectDocument11 pagesMeditation Its Effectmanisami7036Pas encore d'évaluation
- Tracheostomy Tube Brochure - 086034Document4 pagesTracheostomy Tube Brochure - 086034adelin litanPas encore d'évaluation
- Raasn October NewsletterDocument6 pagesRaasn October Newsletterapi-231055558Pas encore d'évaluation
- End-Stage Heart DiseaseDocument44 pagesEnd-Stage Heart DiseaseCyrille AgnesPas encore d'évaluation
- Clinical Guideline For Management of Gallstones Pathology in AdultsDocument11 pagesClinical Guideline For Management of Gallstones Pathology in AdultsrintyosoPas encore d'évaluation
- Transitional CareDocument8 pagesTransitional CareJayvee FerandezPas encore d'évaluation
- Ace ArsenalDocument11 pagesAce ArsenalJustus WeißePas encore d'évaluation
- HIV and PregnancyDocument9 pagesHIV and PregnancyUm HamoOdPas encore d'évaluation
- Hybrid Hues 2008Document196 pagesHybrid Hues 2008Kulsharma100% (1)
- Basics of Pharmacology: Speaker - DR - SantoshDocument76 pagesBasics of Pharmacology: Speaker - DR - SantoshRamya ChallaPas encore d'évaluation
- Bell's Palsy Treatments & Medications - SingleCareDocument11 pagesBell's Palsy Treatments & Medications - SingleCareRoxan PacsayPas encore d'évaluation
- My First Authored PublicationDocument16 pagesMy First Authored PublicationSamuel UzonduPas encore d'évaluation
- Felicias Resume 1Document1 pageFelicias Resume 1api-357153848Pas encore d'évaluation
- Proposal For Defense Group 3Document77 pagesProposal For Defense Group 3Kristiene Kyle AquinoPas encore d'évaluation
- Palpation and Assessment SkillsDocument305 pagesPalpation and Assessment SkillsElin Taopan97% (34)
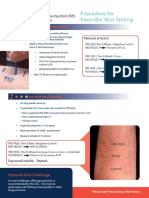
- Procedure For Penicillin Skin TestingDocument4 pagesProcedure For Penicillin Skin TestingHarshan Isuru KumaraPas encore d'évaluation
- HR PWDDocument13 pagesHR PWDDiane UyPas encore d'évaluation
- E B I W: Safemedicate Rounding Rules GuidelinesDocument9 pagesE B I W: Safemedicate Rounding Rules Guidelineslesky17Pas encore d'évaluation
- The PRISMAFLEX System: Making Possible PersonalDocument12 pagesThe PRISMAFLEX System: Making Possible PersonalluisfulaPas encore d'évaluation
- Induction Update2Document30 pagesInduction Update2yusepPas encore d'évaluation
- Hepar Sulph - The Homeopathic AntibioticDocument54 pagesHepar Sulph - The Homeopathic AntibioticGer DersenPas encore d'évaluation
- By - Derina FixDocument1 pageBy - Derina FixSellyta HasugianPas encore d'évaluation
- RetailerPriceList OHDocument1 pageRetailerPriceList OHMuhammad Samiur RahmanPas encore d'évaluation
- MCQ Thoracic SurgeryDocument5 pagesMCQ Thoracic SurgeryKhamis Belal100% (6)
- Nursing Group Research PaperDocument16 pagesNursing Group Research Paperapi-368267454Pas encore d'évaluation
- OT HoppysAppReviewDocument1 pageOT HoppysAppReviewIris De La CalzadaPas encore d'évaluation
- PRICE - LIST - Price List 2023 - Price List Bbraun-DNR Reguler - Basic - Care - 2023Document2 pagesPRICE - LIST - Price List 2023 - Price List Bbraun-DNR Reguler - Basic - Care - 2023yu2nwahyuni.aptPas encore d'évaluation
- Lipoxome Business PresentationDocument20 pagesLipoxome Business PresentationCaerwyn AshPas encore d'évaluation