Vous aimerez peut-être aussi
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (587)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (894)
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (399)
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (73)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5794)
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2219)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (344)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (265)
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (119)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- FasciotomyDocument17 pagesFasciotomyPrasojo JojoPas encore d'évaluation
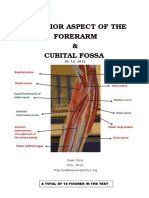
- Anterior Aspect of The Forearm Cubital Fossa 2012Document28 pagesAnterior Aspect of The Forearm Cubital Fossa 2012Elena IftimiPas encore d'évaluation
- Kompartemen SindromDocument9 pagesKompartemen SindromPutri PratiwiPas encore d'évaluation
- Lab 4 - ForearmDocument44 pagesLab 4 - ForearmTayai LengimaPas encore d'évaluation
- Ashrafology's Clinical Examination NotesDocument41 pagesAshrafology's Clinical Examination NotesAnjnaPas encore d'évaluation
- Volkmann's Ischemic ContractureDocument41 pagesVolkmann's Ischemic ContractureAayush AryalPas encore d'évaluation
- Piano Hand TechniqueDocument12 pagesPiano Hand TechniquevaneriolamaPas encore d'évaluation
- Review Notes - Fish & Freiberg Plastic SurgeryDocument24 pagesReview Notes - Fish & Freiberg Plastic SurgeryMartin Susanto, MDPas encore d'évaluation
- Piano Technique - Anatomy of The Hand and Its Relevance To A Fluent Finger TechniqueDocument31 pagesPiano Technique - Anatomy of The Hand and Its Relevance To A Fluent Finger TechniqueMartha Argerich100% (6)
- 9-34 Hand IIIDocument40 pages9-34 Hand IIIMario100% (1)
- Anatomy TopoDocument62 pagesAnatomy TopoDoina GoreanuPas encore d'évaluation
- The Anatomy Lesson of Dr. Nicolaes Tulp, Journal of Hand Surgery (2006)Document10 pagesThe Anatomy Lesson of Dr. Nicolaes Tulp, Journal of Hand Surgery (2006)cjlassPas encore d'évaluation
- Modified Duran Hand Therapy ProtocolDocument4 pagesModified Duran Hand Therapy ProtocolkkPas encore d'évaluation
- One Move Too ManyDocument117 pagesOne Move Too ManyMZ100% (3)
- Examining Peripheral NervesDocument6 pagesExamining Peripheral NervesNur Atiqah ZainalPas encore d'évaluation
- Forelimb Myology of The Golden Pheasant (Chrysolophus Pictus)Document9 pagesForelimb Myology of The Golden Pheasant (Chrysolophus Pictus)sytaPas encore d'évaluation
- Muscles of The Arm and ForearmDocument8 pagesMuscles of The Arm and ForearmالبراءPas encore d'évaluation
- Lab 4 - ForearmDocument54 pagesLab 4 - ForearmmamuuramadanPas encore d'évaluation
- What Can Studying Musicians Tell Us About Motor Control of The Hand?Document16 pagesWhat Can Studying Musicians Tell Us About Motor Control of The Hand?Anonymous ZSmSZErTqtPas encore d'évaluation
- Lies in The Palm of His Hand: A Pianist's SecretDocument8 pagesLies in The Palm of His Hand: A Pianist's SecretDerekHoPas encore d'évaluation
- Muscles of the Forearm CompartmentsDocument88 pagesMuscles of the Forearm Compartmentsromaisa akhtarPas encore d'évaluation
- Muscles of Upper LimbDocument21 pagesMuscles of Upper Limbمحمد علىPas encore d'évaluation
- Front of ForearmDocument19 pagesFront of ForearmAditya AnandPas encore d'évaluation