Vous aimerez peut-être aussi
- Health Benefits of Cascade Fermented Foods - "You'll Just Have To Live With It!" (No Thank You!)Document105 pagesHealth Benefits of Cascade Fermented Foods - "You'll Just Have To Live With It!" (No Thank You!)BiologicalMed99100% (4)
- Med Surg Test 4 Study GuideDocument29 pagesMed Surg Test 4 Study GuideJessPas encore d'évaluation
- Mercury Dental Amalgams: The Controversy Continues by Gary Null, PH.D., and Martin Feldman, M.D.Document55 pagesMercury Dental Amalgams: The Controversy Continues by Gary Null, PH.D., and Martin Feldman, M.D.Gary NullPas encore d'évaluation
- Metabolism of ProteinsDocument50 pagesMetabolism of ProteinsAbdur RehmanPas encore d'évaluation
- Amino Acid Catabolism Between OrgansDocument44 pagesAmino Acid Catabolism Between OrgansFarhati MardhiyahPas encore d'évaluation
- Lectures 1 3 Handout For PrintingDocument43 pagesLectures 1 3 Handout For Printingkriss Wong100% (2)
- Integration of Metabolism Integration of MetabolismDocument10 pagesIntegration of Metabolism Integration of MetabolismEdison LucianoPas encore d'évaluation
- AVS 172 Reproductive Physiology: Amin Ahamdzadeh Department of Animal and Veterinary Science University of IdahoDocument41 pagesAVS 172 Reproductive Physiology: Amin Ahamdzadeh Department of Animal and Veterinary Science University of IdahoCristina CarvalhoPas encore d'évaluation
- HW - Carbohydrate Metabolism II & Lipid MetabolismDocument2 pagesHW - Carbohydrate Metabolism II & Lipid MetabolismyanPas encore d'évaluation
- Lipid Profiles ExplainedDocument5 pagesLipid Profiles ExplainedFahd Abdullah Al-refaiPas encore d'évaluation
- EnzymesDocument21 pagesEnzymesCatherine Merilleno100% (1)
- Cholesterol SynthesisDocument16 pagesCholesterol Synthesispriya19866100% (1)
- Enzymes: Protein Catalysts That Increase The Rate of Reactions Without Being Changed in The Overall ProcessDocument49 pagesEnzymes: Protein Catalysts That Increase The Rate of Reactions Without Being Changed in The Overall ProcessGhafoor AzamPas encore d'évaluation
- Biochemistry of Digestive SystemDocument55 pagesBiochemistry of Digestive SystemSyam UnhasPas encore d'évaluation
- Glycolysis: Shekhar Chandra Yadav Lecturer Dept. of BiochemistryDocument25 pagesGlycolysis: Shekhar Chandra Yadav Lecturer Dept. of BiochemistryTULSI SHARMAPas encore d'évaluation
- Biochemistry Lecture 2 Cell and OrganellesDocument16 pagesBiochemistry Lecture 2 Cell and OrganellesProfessor Rakesh Sharma Biochemistry LecturesPas encore d'évaluation
- Enzymes and Their Importance in Plants and AnimalsDocument4 pagesEnzymes and Their Importance in Plants and Animalsanili50% (2)
- ChromosomesDocument9 pagesChromosomestayyabaPas encore d'évaluation
- 4 Nutrition and Metabolism Lipid-Updated (Compatibility Mode)Document45 pages4 Nutrition and Metabolism Lipid-Updated (Compatibility Mode)Anh Nguyen100% (1)
- Micronutrients in health and disease reviewDocument10 pagesMicronutrients in health and disease reviewLeonard LeonardPas encore d'évaluation
- Carbohydrates Slide-2 PHARM-D, SALUDocument85 pagesCarbohydrates Slide-2 PHARM-D, SALUShahid AhmedPas encore d'évaluation
- Avishkar 2019 PosterDocument1 pageAvishkar 2019 PosterVedant 23 GooglePas encore d'évaluation
- ProteinDocument39 pagesProteinNICHOLE MOJELLO100% (1)
- Endocrine Disruptors and Hormonal CancerDocument8 pagesEndocrine Disruptors and Hormonal CancerPol MaliaPas encore d'évaluation
- The Urea Cycle: A 40-Character SummaryDocument8 pagesThe Urea Cycle: A 40-Character SummaryManohar PattarPas encore d'évaluation
- CARBOHYDRATES Biochem Pre-LabDocument50 pagesCARBOHYDRATES Biochem Pre-LabGlen MangaliPas encore d'évaluation
- Classification of Genetic DiseasesDocument8 pagesClassification of Genetic DiseasesTyn TynPas encore d'évaluation
- Digestive System DisordersDocument11 pagesDigestive System DisordersDanie CabahugPas encore d'évaluation
- Mutations: Causes, Types and Genetic DisordersDocument18 pagesMutations: Causes, Types and Genetic DisordersSushanthPas encore d'évaluation
- Acid-Base Balance and DisodersDocument86 pagesAcid-Base Balance and DisodersPrincewill SeiyefaPas encore d'évaluation
- Reactions of MonosaccharidesDocument15 pagesReactions of Monosaccharidesvishnudurga88% (8)
- ACFrOgB4ugyviBu1XK4Lh1UX8Pt64wVQwa2 Exi6l8nAFhE Uu1QLN5OIKowfbqMbZ48dVUAL2yq7eDi4HBBlOHGoCWigzrxxjl305MoZfTvJxd54XHQjc6yi-YzD8c PDFDocument12 pagesACFrOgB4ugyviBu1XK4Lh1UX8Pt64wVQwa2 Exi6l8nAFhE Uu1QLN5OIKowfbqMbZ48dVUAL2yq7eDi4HBBlOHGoCWigzrxxjl305MoZfTvJxd54XHQjc6yi-YzD8c PDFLpPas encore d'évaluation
- Food HygieneDocument314 pagesFood Hygienefeyisa100% (2)
- Subject: Biochemistry Topic:Lipid Metabolism 2 Lecturer: Dr. Laygo DATE: NOV. 2010Document11 pagesSubject: Biochemistry Topic:Lipid Metabolism 2 Lecturer: Dr. Laygo DATE: NOV. 2010Std DlshsiPas encore d'évaluation
- Hexose Monophosphate Shunt CHEM3119"TITLE"Biometabolism Lecture on HMP Shunt and G6PD DeficiencyDocument18 pagesHexose Monophosphate Shunt CHEM3119"TITLE"Biometabolism Lecture on HMP Shunt and G6PD DeficiencyAbdul Jabbar Abdul JabbarPas encore d'évaluation
- Dna and Rna Powerpoint 2Document46 pagesDna and Rna Powerpoint 2api-267309851Pas encore d'évaluation
- CarbohydratesDocument156 pagesCarbohydrateseiddnewPas encore d'évaluation
- Color Reactions of ProteinsDocument1 pageColor Reactions of ProteinsZarah Pauline JimenezPas encore d'évaluation
- 6.1 Types of NutritionDocument11 pages6.1 Types of NutritionNoor Hidayah SambliPas encore d'évaluation
- Beta OxidationDocument19 pagesBeta Oxidationindra100% (1)
- Clinical Biochemistry - Clinical Pathology and Procedures - VetDocument9 pagesClinical Biochemistry - Clinical Pathology and Procedures - VetHimmet AslanPas encore d'évaluation
- Animal Diseases CollectionDocument94 pagesAnimal Diseases CollectionNabin Kumar YadavPas encore d'évaluation
- Chapter 18 Endocrine SystemDocument40 pagesChapter 18 Endocrine SystemlolasparklePas encore d'évaluation
- Primary Structure of ProteinDocument21 pagesPrimary Structure of ProteinNico RobinPas encore d'évaluation
- 10 Steps Glycolysis ExplainedDocument8 pages10 Steps Glycolysis Explaineddani2703Pas encore d'évaluation
- Protein TherapeuticsDocument14 pagesProtein TherapeuticsSumanth Kumar ReddyPas encore d'évaluation
- Metabolic Pathway of Carbohydrate and GlycolysisDocument22 pagesMetabolic Pathway of Carbohydrate and GlycolysisDarshansinh MahidaPas encore d'évaluation
- Nutrition BiochemDocument45 pagesNutrition BiochemKesha Marie TalloPas encore d'évaluation
- Handouts - CarbohydratesDocument9 pagesHandouts - CarbohydratesJerrold CruzPas encore d'évaluation
- The Adrenal GlandDocument41 pagesThe Adrenal GlandRujha Haniena Ahmad RidzuanPas encore d'évaluation
- Regulation of Gluconeogenesis and Glycogen MetabolismDocument35 pagesRegulation of Gluconeogenesis and Glycogen MetabolismdanielachynaPas encore d'évaluation
- Carbohydrates SummaryDocument9 pagesCarbohydrates SummaryHarold NagunaPas encore d'évaluation
- Endocrine System GuideDocument6 pagesEndocrine System GuideChechan AmbaPas encore d'évaluation
- Biochemistry of Kidneys and UrineDocument18 pagesBiochemistry of Kidneys and UrineAndrias PutriPas encore d'évaluation
- Carbs: The sugars that feed usDocument4 pagesCarbs: The sugars that feed usMaria Sophia Faith100% (1)
- CHEM 140 Unit 8 Lec 1 LipidsDocument93 pagesCHEM 140 Unit 8 Lec 1 LipidsMevil Jane MabrasPas encore d'évaluation
- 1.protein Digestion, Urea Cy, MbbsDocument81 pages1.protein Digestion, Urea Cy, MbbsDebarghya MukherjeePas encore d'évaluation
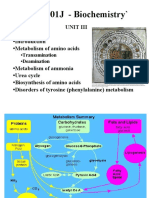
- Biochemistry Unit 03 PDFDocument66 pagesBiochemistry Unit 03 PDFDhanush kannanPas encore d'évaluation
- 15BT103 Biochem-UNIT 3Document53 pages15BT103 Biochem-UNIT 3Adityanair RA1711009010128Pas encore d'évaluation
- Amino Acid Metabolism NotesDocument152 pagesAmino Acid Metabolism NotesAkhilesh TiwariPas encore d'évaluation
- Amino Acid Metabolism NotesDocument8 pagesAmino Acid Metabolism Notessean100% (2)
- Amino Acid Metabolism All LecturesDocument18 pagesAmino Acid Metabolism All Lecturesmizare29gPas encore d'évaluation
- Abdominal X RayDocument64 pagesAbdominal X RayabhishekbmcPas encore d'évaluation
- Plasma Physiology (1-2020) by DR Khaled A AbulfadleDocument9 pagesPlasma Physiology (1-2020) by DR Khaled A AbulfadleUzama Binu AliPas encore d'évaluation
- Digestive System Breakdown and AbsorptionDocument9 pagesDigestive System Breakdown and AbsorptionTana OquendoPas encore d'évaluation
- Tests for Liver Function: Serum BilirubinDocument4 pagesTests for Liver Function: Serum BilirubinHiba EmadPas encore d'évaluation
- Fetal Pig Dissection Lab: Project WeblinkDocument6 pagesFetal Pig Dissection Lab: Project WeblinkJill KoehlerPas encore d'évaluation
- CHRONIC HEPATITIS Prof DR Tarek ShetaDocument38 pagesCHRONIC HEPATITIS Prof DR Tarek ShetaSheren GamaleldenPas encore d'évaluation
- Pathophysiology of Hypovolemic ShockDocument8 pagesPathophysiology of Hypovolemic ShockKAYCEEPas encore d'évaluation
- Cholelytiasis 2013 2015Document37 pagesCholelytiasis 2013 2015Nurul RamadhantyPas encore d'évaluation
- Anatomy of The Digestive Tube of Toco ToucanDocument11 pagesAnatomy of The Digestive Tube of Toco ToucanYinneth SakuritaPas encore d'évaluation
- HepC Handbook Web2008Document151 pagesHepC Handbook Web2008susilorini100% (1)
- Practical 4 Cardivascular and Hepatobiliary SystemDocument9 pagesPractical 4 Cardivascular and Hepatobiliary SystemchinPas encore d'évaluation
- ANEURSYMDocument24 pagesANEURSYMMYLENE GRACE ELARCOSAPas encore d'évaluation
- Mind Map GitDocument7 pagesMind Map Gitronron2008Pas encore d'évaluation
- Liver Alterations in Cats - A Literature ReviewDocument39 pagesLiver Alterations in Cats - A Literature ReviewJoanna ChrysanthouPas encore d'évaluation
- Assessment of Normal DopplerDocument4 pagesAssessment of Normal Dopplerharrinson89Pas encore d'évaluation
- Circulatory System TurtleDocument32 pagesCirculatory System TurtleVillamorchardPas encore d'évaluation
- The Longmire I, II, and III Operations: L. William Traverso, M.D.Document8 pagesThe Longmire I, II, and III Operations: L. William Traverso, M.D.Abu ZidanePas encore d'évaluation
- Innovations and Prospects in Modern Science 13 15.02.23Document481 pagesInnovations and Prospects in Modern Science 13 15.02.23Miroslav Ekman-KremenetskyPas encore d'évaluation
- Pathology Liver Jars FixedDocument7 pagesPathology Liver Jars Fixedapi-3730040Pas encore d'évaluation
- 007-Hepatobiliary & PancreaticDocument59 pages007-Hepatobiliary & PancreaticAhmed ZaghwPas encore d'évaluation
- ABDOMENDocument16 pagesABDOMENHussein Al SaediPas encore d'évaluation
- Cancer: Causes and Risks FactorsDocument88 pagesCancer: Causes and Risks FactorsClancy Anne Garcia NavalPas encore d'évaluation
- AD285230700512 LO230700610: Patient ReportDocument1 pageAD285230700512 LO230700610: Patient ReportReal Gamer 6Pas encore d'évaluation
- Anatomy and Physiology of Biliary TreeDocument48 pagesAnatomy and Physiology of Biliary TreeKamalakanta Das100% (1)
- 2018 Ultrasound A Core Review PDFDocument617 pages2018 Ultrasound A Core Review PDFsun sealPas encore d'évaluation
- Ascites PDFDocument8 pagesAscites PDFJanina Patricia BuddlePas encore d'évaluation
- The Liver in Systemic Disease A Clinician's Guide To Abnormal LiverDocument285 pagesThe Liver in Systemic Disease A Clinician's Guide To Abnormal Liveroleksandra.bilotkachPas encore d'évaluation