Vous aimerez peut-être aussi
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5794)
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (895)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (344)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (399)
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (588)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (266)
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (73)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2259)
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (120)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- Family School Project Lesson Plan AstrologyDocument3 pagesFamily School Project Lesson Plan Astrologyapi-529488210Pas encore d'évaluation
- Epidemiological Triad of HIV/AIDS: AgentDocument8 pagesEpidemiological Triad of HIV/AIDS: AgentRakib HossainPas encore d'évaluation
- Practical CS ProcessingDocument483 pagesPractical CS ProcessinganAMUstudent100% (2)
- Keira Knightley: Jump To Navigation Jump To SearchDocument12 pagesKeira Knightley: Jump To Navigation Jump To SearchCrina LupuPas encore d'évaluation
- Architecture of Neural NWDocument79 pagesArchitecture of Neural NWapi-3798769Pas encore d'évaluation
- Risk Assessment For ExcavationDocument6 pagesRisk Assessment For ExcavationAhmed GamalPas encore d'évaluation
- Lesson Plan Defining and Non Relative Clauses XII (I)Document3 pagesLesson Plan Defining and Non Relative Clauses XII (I)mariaalexeli0% (1)
- Bell Single-Sleeve Shrug Crochet PatternDocument2 pagesBell Single-Sleeve Shrug Crochet PatternsicksoxPas encore d'évaluation
- 2 Beats Per Measure 3 Beats Per Measure 4 Beats Per MeasureDocument24 pages2 Beats Per Measure 3 Beats Per Measure 4 Beats Per MeasureArockiya StephenrajPas encore d'évaluation
- Neuroscience Core ConceptsDocument2 pagesNeuroscience Core Conceptseglantina alishollariPas encore d'évaluation
- Management Glossary - Musa KamawiDocument50 pagesManagement Glossary - Musa KamawiKazi Nazrul IslamPas encore d'évaluation
- SRM 7 EHP 4 Release Notes PDFDocument18 pagesSRM 7 EHP 4 Release Notes PDFMOHAMMED SHEHBAAZPas encore d'évaluation
- Cross System Create Supplier ProcessDocument14 pagesCross System Create Supplier ProcesssakthiroboticPas encore d'évaluation
- Dphs Freshman Course List 2024-25Document1 pageDphs Freshman Course List 2024-25api-337771649Pas encore d'évaluation
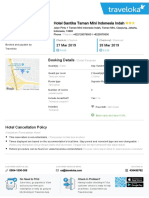
- Hotel Voucher: Itinerary ID Hotel Santika Taman Mini Indonesia IndahDocument2 pagesHotel Voucher: Itinerary ID Hotel Santika Taman Mini Indonesia IndahSyukron PribadiPas encore d'évaluation
- What On Earth Is A MainframeDocument132 pagesWhat On Earth Is A MainframeCarlos DantasPas encore d'évaluation
- Classical Encryption TechniqueDocument18 pagesClassical Encryption TechniquetalebmuhsinPas encore d'évaluation
- RSM222.f22.CourseOutline v3 2022-09-05Document9 pagesRSM222.f22.CourseOutline v3 2022-09-05Kirsten WangPas encore d'évaluation
- QLD Plan Draft Review Raw DataDocument242 pagesQLD Plan Draft Review Raw DataRohit Jain100% (1)
- Module 1Document64 pagesModule 1Jackyson RajkumarPas encore d'évaluation
- Sharp Product-Catalogue 2019 enDocument48 pagesSharp Product-Catalogue 2019 enMiki di KaprioPas encore d'évaluation
- Hybrid Neural-Network - Genetic Algorithm Technique For Aircraft Engine Performance Diagnostics Developed and DemonstratedDocument4 pagesHybrid Neural-Network - Genetic Algorithm Technique For Aircraft Engine Performance Diagnostics Developed and Demonstratedmohamad theibechPas encore d'évaluation
- MetLife CaseDocument4 pagesMetLife Casekatee3847Pas encore d'évaluation
- El Nino 3Document8 pagesEl Nino 3nisAfiqahPas encore d'évaluation
- Education Programme: The School of Concrete TechnologyDocument28 pagesEducation Programme: The School of Concrete TechnologyThabiso Jacob MokwenaPas encore d'évaluation
- Corporate Profile of Multimode GroupDocument6 pagesCorporate Profile of Multimode GroupShaheen RahmanPas encore d'évaluation
- Imcp - RocketbookDocument11 pagesImcp - Rocketbookapi-690398026Pas encore d'évaluation
- DD 3600 3500 3000 Parts CatalogDocument46 pagesDD 3600 3500 3000 Parts CatalogAndres Fdo Mora D100% (2)
- Audit Committee and Corporate Governance: CA Pragnesh Kanabar Sir's The Audit Academy-CA Final AuditDocument17 pagesAudit Committee and Corporate Governance: CA Pragnesh Kanabar Sir's The Audit Academy-CA Final AuditPULKIT MURARKAPas encore d'évaluation
- SET UP Computer ServerDocument3 pagesSET UP Computer ServerRicHArdPas encore d'évaluation