Vous aimerez peut-être aussi
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (119)
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (399)
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (587)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2219)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- 1995 - Burne Hogarth - Dynamic Wrinkles and DraperyDocument145 pages1995 - Burne Hogarth - Dynamic Wrinkles and DraperyLjiljana Antonovic100% (6)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (344)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (890)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (73)
- Bio Finish in TextilesDocument35 pagesBio Finish in Textilessahu_disha100% (2)
- Bakeries and ConfectioneriesDocument26 pagesBakeries and ConfectioneriesMihai UngureanuPas encore d'évaluation
- ) .Mechanical, Chemical, Biological - Moving Towards Closed-Loop Bio-Based - Recycling in A Circular Economy of Sustainable TextilesDocument13 pages) .Mechanical, Chemical, Biological - Moving Towards Closed-Loop Bio-Based - Recycling in A Circular Economy of Sustainable TextilesAydee ChoquePas encore d'évaluation
- CMHEC ManufacturersDocument3 pagesCMHEC ManufacturersTamilarasiPas encore d'évaluation
- Papermaking Raw Materia & Their Characteristics IIDocument115 pagesPapermaking Raw Materia & Their Characteristics IIvivekbhuchemPas encore d'évaluation
- Corporate Bond Market Trends Emerging Risks Monetary PolicyDocument58 pagesCorporate Bond Market Trends Emerging Risks Monetary PolicyRomneck RoveroPas encore d'évaluation
- Cardio VDocument41 pagesCardio VRomneck RoveroPas encore d'évaluation
- Floor Plan With MOH AuthenticationDocument1 pageFloor Plan With MOH AuthenticationRomneck RoveroPas encore d'évaluation
- The Human EyeDocument9 pagesThe Human EyeRomneck RoveroPas encore d'évaluation
- BreastDocument6 pagesBreastRomneck RoveroPas encore d'évaluation
- Meninges and CSF FlowDocument2 pagesMeninges and CSF FlowRomneck RoveroPas encore d'évaluation
- Immune SystemDocument17 pagesImmune SystemRomneck RoveroPas encore d'évaluation
- Introduction to First Aid BasicsDocument13 pagesIntroduction to First Aid BasicsRomneck RoveroPas encore d'évaluation
- Integrated Management of Adolescent and Adult Illnesses AcuteDocument130 pagesIntegrated Management of Adolescent and Adult Illnesses Acutestuffednurse100% (1)
- SocietyDocument26 pagesSocietyRomneck RoveroPas encore d'évaluation
- Cancer Awareness: Jose Mari C. Castro, MD, MPH Medical Specialist I DSWD-NCR April 2010Document6 pagesCancer Awareness: Jose Mari C. Castro, MD, MPH Medical Specialist I DSWD-NCR April 2010Romneck RoveroPas encore d'évaluation
- NursingBulletin IMCIDocument59 pagesNursingBulletin IMCIseigelystic100% (12)
- Congestive Heart FailureDocument35 pagesCongestive Heart FailureRomneck RoveroPas encore d'évaluation
- A Registered NurseDocument1 pageA Registered NurseRomneck RoveroPas encore d'évaluation
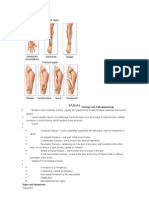
- Etiology Pathophysiology FracturesDocument4 pagesEtiology Pathophysiology FracturesRomneck RoveroPas encore d'évaluation
- LUIDocument2 pagesLUIRomneck RoveroPas encore d'évaluation
- Biology Lecture On ProkaryotesDocument11 pagesBiology Lecture On ProkaryotesRomneck RoveroPas encore d'évaluation
- A Registered NurseDocument1 pageA Registered NurseRomneck RoveroPas encore d'évaluation
- ClaireDocument6 pagesClaireRomneck RoveroPas encore d'évaluation
- AdDocument37 pagesAdAkshat GuptaPas encore d'évaluation
- Peer EffectsDocument54 pagesPeer EffectsRomneck RoveroPas encore d'évaluation
- Toto ResumeDocument2 pagesToto ResumeRomneck RoveroPas encore d'évaluation
- Guidelines For Writing CVDocument2 pagesGuidelines For Writing CVDeody Alvin FranciaPas encore d'évaluation
- Philosophy Is The Study of General and Fundamental Problems Concerning Matters Such As ExistenceDocument2 pagesPhilosophy Is The Study of General and Fundamental Problems Concerning Matters Such As ExistenceRomneck RoveroPas encore d'évaluation
- Biology Lecture On ProkaryotesDocument11 pagesBiology Lecture On ProkaryotesRomneck RoveroPas encore d'évaluation
- United States Patent (10) Patent No.: US 9,375,501 B2Document13 pagesUnited States Patent (10) Patent No.: US 9,375,501 B2Choo Wei shengPas encore d'évaluation
- s13068 019 1529 1 PDFDocument58 pagess13068 019 1529 1 PDFGlan DevadhasPas encore d'évaluation
- 全棉⾯料(100%Cotton Fabric): Item Comp Yarn Weight (G/SM) Width RemarkDocument11 pages全棉⾯料(100%Cotton Fabric): Item Comp Yarn Weight (G/SM) Width RemarkI'm BuddyPas encore d'évaluation
- Reaction Kinetics Glucose - PosterDocument1 pageReaction Kinetics Glucose - PosterZoren A. Del MundoPas encore d'évaluation
- Carbohydrates (Class 4)Document19 pagesCarbohydrates (Class 4)Swetank SahaiPas encore d'évaluation
- Production of 150 Cubic-Meter Bio-Ethanol Per Day From Waste Office PaperDocument119 pagesProduction of 150 Cubic-Meter Bio-Ethanol Per Day From Waste Office PaperPower CellPas encore d'évaluation
- Bio Topic 4 Notes PDFDocument11 pagesBio Topic 4 Notes PDFdgfhjPas encore d'évaluation
- Angelova 2021Document12 pagesAngelova 2021Ritzee MahindrakarPas encore d'évaluation
- Polímeros Biodegradáveis - Uma Solução Parcial para Diminuir A Quantidade Dos Resíduos PlásticosDocument7 pagesPolímeros Biodegradáveis - Uma Solução Parcial para Diminuir A Quantidade Dos Resíduos PlásticosTanara AméliaPas encore d'évaluation
- Eco-Friendly Degumming of Natural Fibers For Textile Applications - A Comprehensive ReviewDocument11 pagesEco-Friendly Degumming of Natural Fibers For Textile Applications - A Comprehensive ReviewWanna MsAbstractPas encore d'évaluation
- Separation of Rayon Thread Thread Using Filter PaperDocument14 pagesSeparation of Rayon Thread Thread Using Filter PaperPandi Selvi ASPas encore d'évaluation
- Bioplastic ProductionDocument2 pagesBioplastic ProductionLukmanKuntajiPas encore d'évaluation
- Food Bioscience: Digambar Kavitake, Sujatha Kandasamy, Palanisamy Bruntha Devi, Prathapkumar Halady ShettyDocument11 pagesFood Bioscience: Digambar Kavitake, Sujatha Kandasamy, Palanisamy Bruntha Devi, Prathapkumar Halady ShettyAlexandruPas encore d'évaluation
- Carbohydrate Polymers: Fedia Bettaieb, Ramzi Khiari, Alain Dufresne, Mohamed Farouk Mhenni, Mohamed Naceur BelgacemDocument6 pagesCarbohydrate Polymers: Fedia Bettaieb, Ramzi Khiari, Alain Dufresne, Mohamed Farouk Mhenni, Mohamed Naceur Belgacemeva diazPas encore d'évaluation
- Journal of Agricultural Science and Technology A PDFDocument15 pagesJournal of Agricultural Science and Technology A PDFyatiPas encore d'évaluation
- Grasim Compre ReportDocument60 pagesGrasim Compre ReportManas KalraPas encore d'évaluation
- Enzyme Application PDFDocument4 pagesEnzyme Application PDFTalib aleemPas encore d'évaluation
- Surface Treatments of Plant Fibers and Their Effects On Mechanical Properties of Fiber-Reinforced Composites: A ReviewDocument16 pagesSurface Treatments of Plant Fibers and Their Effects On Mechanical Properties of Fiber-Reinforced Composites: A Reviewmkumar_234155Pas encore d'évaluation
- Ramie-Degumming Methodologies A Short ReviewDocument7 pagesRamie-Degumming Methodologies A Short ReviewPrabhu TexcomsPas encore d'évaluation
- Advances in Microbial Ecology Volume 8Document315 pagesAdvances in Microbial Ecology Volume 8Fabio OliveiraPas encore d'évaluation
- Topic 4 Prep Questions NewDocument29 pagesTopic 4 Prep Questions NewDIYA SURESHPas encore d'évaluation
- 7 Weingarten2012Document16 pages7 Weingarten2012eze_atn_vPas encore d'évaluation
- Material BahanDocument11 pagesMaterial Bahanimro mahmudahPas encore d'évaluation
- Antimicrobial Silver Nanoparticle Bacterial Cellulose Wound DressingDocument9 pagesAntimicrobial Silver Nanoparticle Bacterial Cellulose Wound Dressingfranciscrick69Pas encore d'évaluation
- Cellulose Crystallinity Index Measurement Techniques and Their Impact On Interpreting Cellulase PerformanceDocument10 pagesCellulose Crystallinity Index Measurement Techniques and Their Impact On Interpreting Cellulase PerformanceParvinder McCartneyPas encore d'évaluation