Vous aimerez peut-être aussi
- Chemistry and Action of Herbicide AntidotesD'EverandChemistry and Action of Herbicide AntidotesFerenc PallosPas encore d'évaluation
- PP 0902000494Document10 pagesPP 090200049410sgPas encore d'évaluation
- (2005) - Suppression of Botrytis Blight of Begonia by Trichoderma Hamatum 382 in Peat and Compost-Amended Potting MixesDocument6 pages(2005) - Suppression of Botrytis Blight of Begonia by Trichoderma Hamatum 382 in Peat and Compost-Amended Potting MixesabdouPas encore d'évaluation
- Howell 2003Document7 pagesHowell 2003jicama SEXTO NIVELPas encore d'évaluation
- 30 36 PDFDocument7 pages30 36 PDFray m deraniaPas encore d'évaluation
- Crop Protectants 2015Document9 pagesCrop Protectants 2015joyeeta8Pas encore d'évaluation
- Phyto 96 0186Document4 pagesPhyto 96 0186KeithSix Yman ArrietaPas encore d'évaluation
- Phyto 03 10 0091Document9 pagesPhyto 03 10 0091Ade PutPas encore d'évaluation
- Anees 2010Document11 pagesAnees 2010Amanda RafaelaPas encore d'évaluation
- Coşkuntuna, Özer - 2008 - Biological Control of Onion Basal Rot Disease Using Trichoderma Harzianum and Induction of Antifungal CompoundDocument7 pagesCoşkuntuna, Özer - 2008 - Biological Control of Onion Basal Rot Disease Using Trichoderma Harzianum and Induction of Antifungal CompoundJosé Matías ZapiolaPas encore d'évaluation
- PBB 4 079Document8 pagesPBB 4 079InnezPas encore d'évaluation
- Trichoderma Species - : Opportunistic, Avirulent Plant SymbiontsDocument14 pagesTrichoderma Species - : Opportunistic, Avirulent Plant SymbiontsSandro ManchegoPas encore d'évaluation
- Won 2003Document14 pagesWon 2003Roberto C. BárcenasPas encore d'évaluation
- Isolation of Trichoderma Spp. and Determination of Their Antifungal, Biochemical and Physiological FeaturesDocument7 pagesIsolation of Trichoderma Spp. and Determination of Their Antifungal, Biochemical and Physiological Featurescrashoverride29187Pas encore d'évaluation
- Isolation of RhizosphereDocument7 pagesIsolation of RhizosphereAnisam AbhiPas encore d'évaluation
- El Debaiky2017 q2Document27 pagesEl Debaiky2017 q2Tammy CevallosPas encore d'évaluation
- Art:10.1007/s11101 006 9032 2Document35 pagesArt:10.1007/s11101 006 9032 2Caroline LopesPas encore d'évaluation
- Tapwal Metode Dual CultureDocument6 pagesTapwal Metode Dual CultureRezki Adi Nugroho 1506114628Pas encore d'évaluation
- Antimicrobial Activity of Trichoderma Harzianum Against Bacteria and FungiDocument8 pagesAntimicrobial Activity of Trichoderma Harzianum Against Bacteria and FungiEmaIžakovićPas encore d'évaluation
- Pseudomonas Fluorescens VSMKU3054 Mediated Induced Systemic ResistanceDocument9 pagesPseudomonas Fluorescens VSMKU3054 Mediated Induced Systemic ResistanceHvzem MohammedPas encore d'évaluation
- Contreras Cornejo 2017Document9 pagesContreras Cornejo 2017Karen Avalos VelaPas encore d'évaluation
- TrichodermaDocument13 pagesTrichodermaAndreina ZamoraPas encore d'évaluation
- Baldotto Et Al, 2011 - Structural Interaction Between GFP Herbaspirillum and PineappleDocument12 pagesBaldotto Et Al, 2011 - Structural Interaction Between GFP Herbaspirillum and PineappleAnita Bonita Bueno NunesPas encore d'évaluation
- Ka Gale 2004Document10 pagesKa Gale 2004hsaddyPas encore d'évaluation
- Physiological and Molecular Plant Pathology: SciencedirectDocument8 pagesPhysiological and Molecular Plant Pathology: SciencedirectGeral93Pas encore d'évaluation
- General Mechanisms of Action of Microbial Biocontrol AgentsDocument12 pagesGeneral Mechanisms of Action of Microbial Biocontrol AgentsMonaia YehiaPas encore d'évaluation
- Ebpc D 22 00026Document18 pagesEbpc D 22 00026Ayman AhmedPas encore d'évaluation
- Inhibition of in Vitro Growth of Soil-Borne Pathogens by Compost-Inhabiting Indigenous Bacteria and FungiDocument7 pagesInhibition of in Vitro Growth of Soil-Borne Pathogens by Compost-Inhabiting Indigenous Bacteria and FungiPetruta Calina CorneaPas encore d'évaluation
- Antifungal Activity Against Plant Pathogenic Fungi of Chaetoviridins Isolated From Chaetomium GlobosumDocument5 pagesAntifungal Activity Against Plant Pathogenic Fungi of Chaetoviridins Isolated From Chaetomium GlobosummaczweiPas encore d'évaluation
- 2022 - Priming of Defense Related Genes in Brassica Oleracea Var. CapitataDocument10 pages2022 - Priming of Defense Related Genes in Brassica Oleracea Var. CapitataMariana Rocha MaximianoPas encore d'évaluation
- Phyto 08 10 0224Document13 pagesPhyto 08 10 0224Jose Francisco Diaz NajeraPas encore d'évaluation
- Biocontrol PDFDocument6 pagesBiocontrol PDFWesh De GeminosPas encore d'évaluation
- Rista Susanti - Plant Ecophysiology - Plant Response To Microorganisms Systemic Immunity Triggered by Local Plant Microbe InteractionDocument7 pagesRista Susanti - Plant Ecophysiology - Plant Response To Microorganisms Systemic Immunity Triggered by Local Plant Microbe InteractionRISTA SUSANTIPas encore d'évaluation
- Mejora Del Antagosnismo y Tolerancia A Los Fungicidas en Aislados de Tricoderma Con UVDocument9 pagesMejora Del Antagosnismo y Tolerancia A Los Fungicidas en Aislados de Tricoderma Con UVJalcamPas encore d'évaluation
- 5 Asadetal - PJM 2014pdfDocument11 pages5 Asadetal - PJM 2014pdfAgung Wiriat PutraPas encore d'évaluation
- University of Horticultural Science, Bagalkot College of Horticulture, Bagalkot, 587104Document39 pagesUniversity of Horticultural Science, Bagalkot College of Horticulture, Bagalkot, 587104Arjunsinh RathavaPas encore d'évaluation
- Rhizosphere Community Selection Reveals Bacteria Associated With Reduced Root DiseaseDocument27 pagesRhizosphere Community Selection Reveals Bacteria Associated With Reduced Root DiseaseAbdul NasirPas encore d'évaluation
- Sciencedirect: Biotic and Abiotic Stress-Responsive Genes Are Stimulated To Resist Drought Stress in Purple WheatDocument18 pagesSciencedirect: Biotic and Abiotic Stress-Responsive Genes Are Stimulated To Resist Drought Stress in Purple WheatChrisYapPas encore d'évaluation
- EurJourPlPathol2008 PrimingDocument10 pagesEurJourPlPathol2008 PrimingPatrícia JoãoPas encore d'évaluation
- Production of Enzymes and Phytotoxins by The Plant PathogenDocument40 pagesProduction of Enzymes and Phytotoxins by The Plant PathogenMudit MisraPas encore d'évaluation
- The Effect of Plant Extracts As Seed Treatments To Control Bacterial Leaf Spot of Tomato in TanzaniaDocument10 pagesThe Effect of Plant Extracts As Seed Treatments To Control Bacterial Leaf Spot of Tomato in TanzaniaAdita Nur FaoziyahPas encore d'évaluation
- Collectotrichum Acutatum: Sookyeong Hwang, Joohyung Kim, Young Ho Kim and Heung Tae KimDocument7 pagesCollectotrichum Acutatum: Sookyeong Hwang, Joohyung Kim, Young Ho Kim and Heung Tae KimMun Shan KohPas encore d'évaluation
- 060 JimenezDocument6 pages060 JimenezAri Puji AstutiPas encore d'évaluation
- Powdery Mildew 2008 - Genetics and ResistanceDocument5 pagesPowdery Mildew 2008 - Genetics and ResistancenormanwillowPas encore d'évaluation
- Prospect of Trichoderma As A Potential FungicideDocument10 pagesProspect of Trichoderma As A Potential FungicideInternational Journal of Human Genetics Medical Biotechnology & Microbiological StudiesPas encore d'évaluation
- Fritz2006 PDFDocument7 pagesFritz2006 PDFMarichuAlvaradoPas encore d'évaluation
- Abu-Nada Et Al 2007Document17 pagesAbu-Nada Et Al 2007Keyla GonzálezPas encore d'évaluation
- 2015 Article 327Document11 pages2015 Article 327Dicson Sánchez AbadPas encore d'évaluation
- PhytoalexinsDocument6 pagesPhytoalexinsNarendra NaniPas encore d'évaluation
- Plant Pathogen: Agrobacterium Is Well Known For Its Ability To TransferDocument4 pagesPlant Pathogen: Agrobacterium Is Well Known For Its Ability To Transfergupta_archana9Pas encore d'évaluation
- Tmp275e TMPDocument5 pagesTmp275e TMPFrontiersPas encore d'évaluation
- TRi Sahebani2008 PDFDocument5 pagesTRi Sahebani2008 PDFChi Đoàn TtutPas encore d'évaluation
- Current Research and Innovations in Plant Pathology Chapter 4 Host Defence Mechanisms in Plant DiseasesDocument18 pagesCurrent Research and Innovations in Plant Pathology Chapter 4 Host Defence Mechanisms in Plant DiseasesRajat SharmaPas encore d'évaluation
- Plant Pathogenicity Phenotyping of Ralstonia Solanacearum StrainsDocument17 pagesPlant Pathogenicity Phenotyping of Ralstonia Solanacearum StrainsAlberto MachoPas encore d'évaluation
- tmpA2A1 TMPDocument2 pagestmpA2A1 TMPFrontiersPas encore d'évaluation
- 8954-Texto Del Artículo-46334-2-10-20190310Document7 pages8954-Texto Del Artículo-46334-2-10-20190310Jorge TurpoPas encore d'évaluation
- TMP 3 F59Document2 pagesTMP 3 F59FrontiersPas encore d'évaluation
- Regulation of Enteric Endophytic Bacterial Colonization by Plant DefensesDocument10 pagesRegulation of Enteric Endophytic Bacterial Colonization by Plant DefensesDiegoPas encore d'évaluation
- Baysal, Çalışkan, Yeşilova - 2008 - An Inhibitory Effect of A New Bacillus Subtilis Strain (EU07) Against Fusarium Oxysporum F. Sp. Radi PDFDocument8 pagesBaysal, Çalışkan, Yeşilova - 2008 - An Inhibitory Effect of A New Bacillus Subtilis Strain (EU07) Against Fusarium Oxysporum F. Sp. Radi PDFJosé Matías ZapiolaPas encore d'évaluation
- Phage InfectionDocument8 pagesPhage InfectionMahmoud GaberPas encore d'évaluation
- The Adventures of Sherlock HolmesDocument368 pagesThe Adventures of Sherlock HolmesUtanka DePas encore d'évaluation
- Pdis 1999 83 11 972 PDFDocument12 pagesPdis 1999 83 11 972 PDFUtanka DePas encore d'évaluation
- Pdis 1998 82 5 501 PDFDocument6 pagesPdis 1998 82 5 501 PDFUtanka DePas encore d'évaluation
- Special Issue: Trichoderma - From Basic Biology To BiotechnologyDocument2 pagesSpecial Issue: Trichoderma - From Basic Biology To BiotechnologyUtanka DePas encore d'évaluation
- Review On PGPR Mechanism 2012 PDFDocument16 pagesReview On PGPR Mechanism 2012 PDFUtanka DePas encore d'évaluation
- Brundrett 2002 Evolution Mycorrhiza Fungi Roots Plants PDFDocument31 pagesBrundrett 2002 Evolution Mycorrhiza Fungi Roots Plants PDFUtanka DePas encore d'évaluation
- Brundrett 2002 Evolution Mycorrhiza Fungi Roots Plants PDFDocument31 pagesBrundrett 2002 Evolution Mycorrhiza Fungi Roots Plants PDFUtanka DePas encore d'évaluation
- Multiple MyelomaDocument47 pagesMultiple MyelomaUtanka De100% (6)
- Professor Asish De: Introductory Remarks and Welcome AddressDocument5 pagesProfessor Asish De: Introductory Remarks and Welcome AddressUtanka DePas encore d'évaluation
- Phylogenetic Distribution and Evolution of MycorrhizasDocument65 pagesPhylogenetic Distribution and Evolution of MycorrhizasUtanka DePas encore d'évaluation
- 6 Malt Extract Agar BrothDocument2 pages6 Malt Extract Agar BrothUtanka DePas encore d'évaluation
- Candidiasis: by Muhammad-Amin Shamall Muhammad-Amin Supervised by Dr. Hero IsmealDocument10 pagesCandidiasis: by Muhammad-Amin Shamall Muhammad-Amin Supervised by Dr. Hero IsmealBunny OzuPas encore d'évaluation
- Atrium Symposium LINC 2012 Dr. GoverdeDocument45 pagesAtrium Symposium LINC 2012 Dr. GoverdegodefroyciosiPas encore d'évaluation
- Hain GenoType Line Probe Assay TestDocument20 pagesHain GenoType Line Probe Assay Testpacome abdulPas encore d'évaluation
- Research Paper Herbal MedicineDocument4 pagesResearch Paper Herbal Medicineaflbtlzaw100% (1)
- Journal Neuropediatri PDFDocument8 pagesJournal Neuropediatri PDFHalimah PramudiyantiPas encore d'évaluation
- Clinical Study: Overjet and Overbite Influence On Cyclic Masticatory Movements: A CT StudyDocument7 pagesClinical Study: Overjet and Overbite Influence On Cyclic Masticatory Movements: A CT Studyarya091193Pas encore d'évaluation
- Dental ImplantsDocument1 pageDental ImplantsPaWan VelagaPas encore d'évaluation
- Bipolar Disorder Journal Reading (International Journal Research of Pharmacy)Document19 pagesBipolar Disorder Journal Reading (International Journal Research of Pharmacy)Imron RosyadiPas encore d'évaluation
- Practice Guidelines For Obstetric Anesthesia An Updated Report by The American Society of Anesthesiologists Task Force On Obstetric AnesthesiaDocument14 pagesPractice Guidelines For Obstetric Anesthesia An Updated Report by The American Society of Anesthesiologists Task Force On Obstetric AnesthesiaMadalina TalpauPas encore d'évaluation
- Acute Purulent Diseases of Fingers and HandDocument23 pagesAcute Purulent Diseases of Fingers and Handluckyswiss7776848Pas encore d'évaluation
- Stroke Signs and Symptoms - CDC - GovDocument2 pagesStroke Signs and Symptoms - CDC - Govjeanne_mbPas encore d'évaluation
- Head Trauma Brain InjuryDocument54 pagesHead Trauma Brain Injurymirzabb1Pas encore d'évaluation
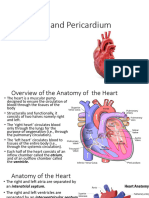
- The Heart and PericardiumDocument40 pagesThe Heart and PericardiumMartha Orendu Oche AttahPas encore d'évaluation
- History and Physical Exam GuidelinesDocument5 pagesHistory and Physical Exam GuidelinesPrincella MonicaPas encore d'évaluation
- Kidney Failure ThesisDocument8 pagesKidney Failure Thesisafcngxbbx100% (3)
- Automated CLIA Analyzers-171222 - ITKDocument14 pagesAutomated CLIA Analyzers-171222 - ITKmPas encore d'évaluation
- RS3772 - Grade - 9 Principle and Practices of Fruit Crop ProductionDocument96 pagesRS3772 - Grade - 9 Principle and Practices of Fruit Crop ProductionMithlesh YadavPas encore d'évaluation
- IM-Module B Summarized Notes (IBD)Document69 pagesIM-Module B Summarized Notes (IBD)DeepbluexPas encore d'évaluation
- MCQ Biochemistry LipidsDocument38 pagesMCQ Biochemistry LipidsstuffPas encore d'évaluation
- Anesthesia For in Vitro Fertilization: Review ArticleDocument11 pagesAnesthesia For in Vitro Fertilization: Review ArticleSumangala MagiPas encore d'évaluation
- Nur 102Document3 pagesNur 102Fretchel Grace Silverado MesaPas encore d'évaluation
- Laboratory Tests InterpretationDocument12 pagesLaboratory Tests InterpretationKaloy Kamao100% (1)
- Top 10 Differential Diagnoses in Family Medicine - CoughDocument2 pagesTop 10 Differential Diagnoses in Family Medicine - CoughTiza OyonPas encore d'évaluation
- Binomial Distribution ExercisesDocument3 pagesBinomial Distribution ExercisesAlp Eren AKYUZPas encore d'évaluation
- 5 Nursing Services Scheduling PolicyDocument7 pages5 Nursing Services Scheduling PolicyVin BitzPas encore d'évaluation
- Logbookgazett MOHDocument56 pagesLogbookgazett MOHShima OnnPas encore d'évaluation
- LCB Surgical Technique - 2022 Update v3Document24 pagesLCB Surgical Technique - 2022 Update v3Samuel SalvadorPas encore d'évaluation
- 2018 Chapter 3-1 2018 Introduction To Industrial Higiene & Occupational Health PDFDocument59 pages2018 Chapter 3-1 2018 Introduction To Industrial Higiene & Occupational Health PDFLove StrikePas encore d'évaluation
- NRC Guidelines GujrathDocument68 pagesNRC Guidelines GujrathNirajkumar KondekarPas encore d'évaluation
- Bulacan State UniversityDocument5 pagesBulacan State UniversityAlexandra CamuaPas encore d'évaluation