Vous aimerez peut-être aussi
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (119)
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (399)
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (587)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2219)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- PLR CourseDocument43 pagesPLR CourseTaniya ChatterjeePas encore d'évaluation
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (344)
- Tai Chi Qigong TrainerDocument11 pagesTai Chi Qigong Trainerskycall28Pas encore d'évaluation
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (890)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- 6th Central Pay Commission Salary CalculatorDocument15 pages6th Central Pay Commission Salary Calculatorrakhonde100% (436)
- 6th Central Pay Commission Salary CalculatorDocument15 pages6th Central Pay Commission Salary Calculatorrakhonde100% (436)
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (73)
- Infectious DiseasesDocument4 pagesInfectious Diseasesapi-484776271Pas encore d'évaluation
- RMT TrainningDocument2 pagesRMT TrainningDita Nadya RizkitaPas encore d'évaluation
- Intelligent Dosing of RonozymeDocument25 pagesIntelligent Dosing of RonozymeRaymundus Genty Laras100% (1)
- Fradeani User GuideDocument9 pagesFradeani User GuideNador AbdennourPas encore d'évaluation
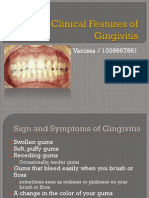
- Clinical Features of GingivitisDocument26 pagesClinical Features of GingivitisVanissa KarisPas encore d'évaluation
- SW 103 2nd Topic PDFDocument12 pagesSW 103 2nd Topic PDFMarielle BringasPas encore d'évaluation
- Manufacturer Qualification in JakartaDocument15 pagesManufacturer Qualification in JakartaMillicent SongsterPas encore d'évaluation
- Brady J. - Environmental Management in Organizations. The IEMA Handbook (2005)Document459 pagesBrady J. - Environmental Management in Organizations. The IEMA Handbook (2005)Wan Naqib Hamidon100% (5)
- Faq BiDocument2 pagesFaq BiShuhaidah SalehinPas encore d'évaluation
- Germplasm Utilization 7Document8 pagesGermplasm Utilization 7Shuhaidah SalehinPas encore d'évaluation
- A Genome-Wide Association Study Identifies ITGA9Document6 pagesA Genome-Wide Association Study Identifies ITGA9Shuhaidah SalehinPas encore d'évaluation
- Fonterra Brands (Malaysia) SDN BHDDocument2 pagesFonterra Brands (Malaysia) SDN BHDShuhaidah Salehin0% (1)
- Banana Housekeeping Reference GenesDocument14 pagesBanana Housekeeping Reference GenesShuhaidah SalehinPas encore d'évaluation
- Banana Housekeeping Reference GenesDocument14 pagesBanana Housekeeping Reference GenesShuhaidah SalehinPas encore d'évaluation
- Journal 5Document8 pagesJournal 5Shuhaidah SalehinPas encore d'évaluation
- 2012 Article 9631Document14 pages2012 Article 9631Shuhaidah SalehinPas encore d'évaluation
- Journal 6Document9 pagesJournal 6Shuhaidah SalehinPas encore d'évaluation
- Journal 2Document9 pagesJournal 2Shuhaidah SalehinPas encore d'évaluation
- NF-kB Inhibitor ACA Shows Activity Against Multiple MyelomaDocument8 pagesNF-kB Inhibitor ACA Shows Activity Against Multiple MyelomaShuhaidah SalehinPas encore d'évaluation
- Journal 3Document12 pagesJournal 3Shuhaidah SalehinPas encore d'évaluation
- Nzgrassland Publication 171Document4 pagesNzgrassland Publication 171Shuhaidah SalehinPas encore d'évaluation
- Asbmt: BK Virus Disease After Allogeneic Stem Cell Transplantation: A Cohort AnalysisDocument7 pagesAsbmt: BK Virus Disease After Allogeneic Stem Cell Transplantation: A Cohort Analysiscalixto hernandezPas encore d'évaluation
- Gross Pulmonary Lesions of Bovine Lung Slaughtered at Jimma Municipality Abattoir, EthiopiaDocument3 pagesGross Pulmonary Lesions of Bovine Lung Slaughtered at Jimma Municipality Abattoir, EthiopiaSolomonPas encore d'évaluation
- Influence of Academic Work on SHS Students' Mental HealthDocument67 pagesInfluence of Academic Work on SHS Students' Mental HealthAliekfarsey Masukat100% (1)
- Silabus Semeste 4Document2 pagesSilabus Semeste 4Widya TumanggorPas encore d'évaluation
- Synopsis of Smart Health PredictionDocument22 pagesSynopsis of Smart Health PredictionSujeet Kumar50% (4)
- HezekiahWalters Plans 3Document34 pagesHezekiahWalters Plans 3Chris VaughnPas encore d'évaluation
- Fragrance ReportDocument44 pagesFragrance ReportIani MarinescuPas encore d'évaluation
- Peace Corps Violation of Office of Victim Advocate Clearances by OVA Employee DI-16-0254 WB CommentsDocument26 pagesPeace Corps Violation of Office of Victim Advocate Clearances by OVA Employee DI-16-0254 WB CommentsAccessible Journal Media: Peace Corps DocumentsPas encore d'évaluation
- Name: Jaanvi Mahajan Course: 2ballb Hons. REGISTRATION NO.: 22212033 Subject: Law and Medicine Submitted To: Mrs. Vijaishree Dubey PandeyDocument9 pagesName: Jaanvi Mahajan Course: 2ballb Hons. REGISTRATION NO.: 22212033 Subject: Law and Medicine Submitted To: Mrs. Vijaishree Dubey Pandeyjaanvi mahajanPas encore d'évaluation
- Hospital: TotalDocument82 pagesHospital: TotalStanford LeePas encore d'évaluation
- Ujjwala Saga PDFDocument84 pagesUjjwala Saga PDFRishi ModiPas encore d'évaluation
- 2015 EHS Challanges Enablon Reportv9Document15 pages2015 EHS Challanges Enablon Reportv9Anonymous sfY8T3q0100% (1)
- DelegationDocument24 pagesDelegationJP FernandezPas encore d'évaluation
- User Manual: Listening To LifeDocument12 pagesUser Manual: Listening To LifeVitor FilipePas encore d'évaluation
- Liver Disease: Overview of The LiverDocument6 pagesLiver Disease: Overview of The LiverMustafa AlmasoudiPas encore d'évaluation
- Classic Series Price List Effective February 2023Document1 pageClassic Series Price List Effective February 2023Yusuf Patria SandiPas encore d'évaluation
- New Improved CV 2018 MayDocument4 pagesNew Improved CV 2018 MayGiorgos Allen WattsPas encore d'évaluation
- Providing Public Area ServicesDocument48 pagesProviding Public Area ServicesFONCY BUYANPas encore d'évaluation
- Kriya Sharir Syllabus 1st Year BamsDocument5 pagesKriya Sharir Syllabus 1st Year BamsHArshil KumArPas encore d'évaluation
- Lab-Ncm 105-Learn Mat Topic 1 WC WL Ob 2022-2023 1st SemDocument6 pagesLab-Ncm 105-Learn Mat Topic 1 WC WL Ob 2022-2023 1st SemDobby AsahiPas encore d'évaluation