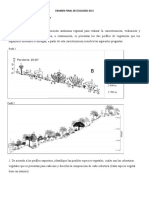
Vous aimerez peut-être aussi
- Encinos. Respuestas Morfofisiológicas a la sequíaD'EverandEncinos. Respuestas Morfofisiológicas a la sequíaPas encore d'évaluation
- Libélulas y caballitos del diablo del departamento del Meta, ColombiaD'EverandLibélulas y caballitos del diablo del departamento del Meta, ColombiaPas encore d'évaluation
- Movilidad de Aves Entre Fragmentos de BosqueDocument8 pagesMovilidad de Aves Entre Fragmentos de BosquecrisanpeloPas encore d'évaluation
- Insectos e Infestaciones Asociadas Al Follaje de Nothofagus AntarcticaDocument11 pagesInsectos e Infestaciones Asociadas Al Follaje de Nothofagus AntarcticamkalkynPas encore d'évaluation
- 2222Document2 pages2222thelasth3r03Pas encore d'évaluation
- Nates-Parra G Et Al. 2008. Efecto Del Cambio Del Paisaje en La Estructura de La Comunidad e Abejas Sin Aguijón en Meta, Colombia PDFDocument14 pagesNates-Parra G Et Al. 2008. Efecto Del Cambio Del Paisaje en La Estructura de La Comunidad e Abejas Sin Aguijón en Meta, Colombia PDFLexy Paola Aristizábal OsorioPas encore d'évaluation
- Gbernardello,+Journal+Manager,+09 DelgadoDocument20 pagesGbernardello,+Journal+Manager,+09 Delgadoyeyi02Pas encore d'évaluation
- Ectatommas en El Cerrejón Como IndicadorDocument11 pagesEctatommas en El Cerrejón Como IndicadorJuan David LozanoPas encore d'évaluation
- ARTICULO WaldemaralegriaDocument25 pagesARTICULO WaldemaralegriawalquerxPas encore d'évaluation
- Efecto de La Fragmentación en La Composición y Estructura Del Bosque Tropical Lluvioso en MéxicoDocument18 pagesEfecto de La Fragmentación en La Composición y Estructura Del Bosque Tropical Lluvioso en MéxicoSalvador Mandujano RodríguezPas encore d'évaluation
- Fontalvo Rodríguez&Domínguez HaydarY.2009Document11 pagesFontalvo Rodríguez&Domínguez HaydarY.2009Yamileth Dominguez-HaydarPas encore d'évaluation
- 1 PBDocument10 pages1 PBcristopher jimenez orozcoPas encore d'évaluation
- Características Edáficas y Composición Florística Del Bosque Estacionalmente Seco La Menta y Timbes, Región Piura, PerúDocument14 pagesCaracterísticas Edáficas y Composición Florística Del Bosque Estacionalmente Seco La Menta y Timbes, Región Piura, Perúpashkito2014Pas encore d'évaluation
- Diversidad Cali ForestalDocument19 pagesDiversidad Cali ForestalJuan Fernando RestrepoPas encore d'évaluation
- Identificación y Caracterización DendrológicaDocument56 pagesIdentificación y Caracterización DendrológicaRom Vargas100% (1)
- Estudio Poblacional de La Palma Camedor en Cuatro Áreas Naturales ProtegidasDocument55 pagesEstudio Poblacional de La Palma Camedor en Cuatro Áreas Naturales ProtegidasMigue Angel Perez Farrera0% (1)
- Prosopis Flexuosa DC. Fabaceae MimosoideaeDocument15 pagesProsopis Flexuosa DC. Fabaceae MimosoideaeEduardo Antonio Molinari NovoaPas encore d'évaluation
- Dialnet RecuperacionDeUnEcosistemaPerturbadoEnElCerroElCas 6140334 PDFDocument8 pagesDialnet RecuperacionDeUnEcosistemaPerturbadoEnElCerroElCas 6140334 PDFBelén HernándezPas encore d'évaluation
- Ruiz y Aguirre. Perija. ColombiaDocument14 pagesRuiz y Aguirre. Perija. ColombiaCesar RuizPas encore d'évaluation
- Avifauna Asociada A Bosques Primarios yDocument18 pagesAvifauna Asociada A Bosques Primarios yLeonărdo Æ. Alcántar RodríguezPas encore d'évaluation
- Art IndividualDocument14 pagesArt IndividualandreaPas encore d'évaluation
- Quispe JuanDocument27 pagesQuispe JuanJuan Pablo Quispe CastilloPas encore d'évaluation
- Especies Del Bosque SubandinoDocument3 pagesEspecies Del Bosque SubandinodarlellyPas encore d'évaluation
- Epifitas Vasculares Bromelias y OrquideasDocument16 pagesEpifitas Vasculares Bromelias y OrquideasMili Gonzales TorresPas encore d'évaluation
- Biografia LeslieHoldridgeDocument6 pagesBiografia LeslieHoldridgewuariPas encore d'évaluation
- 14Document8 pages14Andres Alejandro CamargoPas encore d'évaluation
- 2001 Galemis 13 63Document16 pages2001 Galemis 13 63Joel CordovaPas encore d'évaluation
- Ortiz Pulido2000Document16 pagesOrtiz Pulido2000EDNA CAROLINA SANCHEZ CHAVEZPas encore d'évaluation
- COLECCIÓN DE MACROMICETES VERSION ADRI - Version PreliminarDocument34 pagesCOLECCIÓN DE MACROMICETES VERSION ADRI - Version PreliminarAdriana CorralesPas encore d'évaluation
- El Hotspot Chileno, Prioridad Mundial para La ConservaciónDocument3 pagesEl Hotspot Chileno, Prioridad Mundial para La ConservaciónTiene FloresPas encore d'évaluation
- Fauna de Mil PiesDocument25 pagesFauna de Mil PiesChristian Campos AlvaradoPas encore d'évaluation
- Aves y Mamiferos Del Bosque PDFDocument28 pagesAves y Mamiferos Del Bosque PDFJulio San Martín ÓrdenesPas encore d'évaluation
- Diversidad de Grupos Funcionales de Plantas Del Dosel de Un Bosque Húmedo Tropical Del Alto Orinoco, Estado Amazonas, VenezuelaDocument16 pagesDiversidad de Grupos Funcionales de Plantas Del Dosel de Un Bosque Húmedo Tropical Del Alto Orinoco, Estado Amazonas, Venezuelalindys mirandaPas encore d'évaluation
- 2007 8706 Rmbiodiv 89 01 134Document15 pages2007 8706 Rmbiodiv 89 01 134David MoralesPas encore d'évaluation
- Aguirre 2003 Composición Floristica, Endemismo y Etnobotanica de La Vegetación Del SectorDocument9 pagesAguirre 2003 Composición Floristica, Endemismo y Etnobotanica de La Vegetación Del SectorVanesa Ramos AbensurPas encore d'évaluation
- Parcial de Ecologia AplicadaDocument18 pagesParcial de Ecologia AplicadaJHON JAIRO PARRADO CONTRERASPas encore d'évaluation
- Composicion Floristica de Dos BosquesDocument33 pagesComposicion Floristica de Dos BosquesLewisKarlPas encore d'évaluation
- Manual de Métodos y Criterios para La Evaluación y Monitoreo de La Flora y La VegetaciónDocument23 pagesManual de Métodos y Criterios para La Evaluación y Monitoreo de La Flora y La VegetaciónAndrés YotvelPas encore d'évaluation
- Mariposas Atlantico Boletin CaldasDocument17 pagesMariposas Atlantico Boletin CaldasFabricio Murrieta ChotaPas encore d'évaluation
- Bosque Seco Tropical en La Región CaribeDocument3 pagesBosque Seco Tropical en La Región Caribeangelica villaloboPas encore d'évaluation
- Evaluación de Hongos Ectomicorrícicos Del Grupo de Los Basidiomicetos en La Zona Del Cofre de Perote, VeracruzDocument10 pagesEvaluación de Hongos Ectomicorrícicos Del Grupo de Los Basidiomicetos en La Zona Del Cofre de Perote, VeracruzLuisDomínguezPas encore d'évaluation
- Articulo de Conservacion El ManantialDocument14 pagesArticulo de Conservacion El ManantialMario VargasPas encore d'évaluation
- Bases Ecologicas FinalDocument20 pagesBases Ecologicas FinalGerson Yauli PalominoPas encore d'évaluation
- 2016-Murillo-Ramos Et Al.-Ecologia AustralDocument10 pages2016-Murillo-Ramos Et Al.-Ecologia AustralGylbert RiveraPas encore d'évaluation
- Especies Leñosas Útiles de La Selva Baja Caducifolia en La Sierra de Nanchititla, MéxicoDocument19 pagesEspecies Leñosas Útiles de La Selva Baja Caducifolia en La Sierra de Nanchititla, MéxicoLuisDomínguezPas encore d'évaluation
- 2 Brito Et Al 2014 81 9 16Document8 pages2 Brito Et Al 2014 81 9 16Alice Cortez GavilanesPas encore d'évaluation
- Clave para Identificar ArañasDocument19 pagesClave para Identificar ArañasNicole PinzónPas encore d'évaluation
- Hon - Riqueza y Diversidad de Las Comunidades de Aves Depto. de Francisco Morazán HondurasDocument14 pagesHon - Riqueza y Diversidad de Las Comunidades de Aves Depto. de Francisco Morazán HondurasEdwar H GuarínPas encore d'évaluation
- Gremios EcologicosDocument10 pagesGremios EcologicosVictor Joseph Amasifuen Oroche100% (1)
- Tipos de Vegetacion Mexico PDFDocument15 pagesTipos de Vegetacion Mexico PDFBrayan QuiñoneszPas encore d'évaluation
- Gremios Ecológicos PDFDocument10 pagesGremios Ecológicos PDFAlfred HernanPas encore d'évaluation
- Abundancia Relativa Zorra GrisDocument15 pagesAbundancia Relativa Zorra GrisRoberto Rivera GuzmanPas encore d'évaluation
- V19n1a02 PDFDocument18 pagesV19n1a02 PDFMeriele Rebolledo ContrerasPas encore d'évaluation
- Caracterizacionfloristica BSTTasajeroDocument20 pagesCaracterizacionfloristica BSTTasajeroMiller FernandezPas encore d'évaluation
- UNIVERSIDAD TÉCNICA PARTICULAR DE LOJA Proyecto Botanica RectificadoahoraDocument10 pagesUNIVERSIDAD TÉCNICA PARTICULAR DE LOJA Proyecto Botanica RectificadoahoraSarah NguyenPas encore d'évaluation
- Plantas Del Bosque AndinoDocument12 pagesPlantas Del Bosque AndinoFredy QuinteroPas encore d'évaluation
- Clase 1 de Desarrollo Sustentable Unidad DosDocument37 pagesClase 1 de Desarrollo Sustentable Unidad DosJuana GómezPas encore d'évaluation
- Determinación Del Tamaño Poblacional de La TapirDocument9 pagesDeterminación Del Tamaño Poblacional de La TapiryeniferPas encore d'évaluation
- Bissectus (Vell.) Harling A LA BROTACIÓNDocument8 pagesBissectus (Vell.) Harling A LA BROTACIÓNJoe Saldaña RojasPas encore d'évaluation
- Árboles maderables de la península de Osa: Madera y cortezaD'EverandÁrboles maderables de la península de Osa: Madera y cortezaPas encore d'évaluation
- Métodos de Conservación y Restauración de Materiales Metálicos de Interés HistóricoDocument3 pagesMétodos de Conservación y Restauración de Materiales Metálicos de Interés HistóricokarenciPas encore d'évaluation
- Equipamiento Campamento de MontanaDocument3 pagesEquipamiento Campamento de Montanajoaquin canaviriPas encore d'évaluation
- Examen BienesDocument18 pagesExamen BienesMARIA Calderon sierraPas encore d'évaluation
- Dieta Post Cirugía ProctológicaDocument3 pagesDieta Post Cirugía ProctológicaAndrés OsorioPas encore d'évaluation
- TP Laboratorio - CalorimetríaDocument6 pagesTP Laboratorio - CalorimetríaMauricioPas encore d'évaluation
- Creencias de ItaliaDocument1 pageCreencias de ItaliaYoutube RMPas encore d'évaluation
- Ensayo de Importancia Del Uso de La Fisica en FisioterapiaDocument8 pagesEnsayo de Importancia Del Uso de La Fisica en Fisioterapiaolgaph86Pas encore d'évaluation
- Oxidación de Los Ácidos GrasosDocument2 pagesOxidación de Los Ácidos GrasosPERFIL031Pas encore d'évaluation
- Como Programar Un PLCDocument173 pagesComo Programar Un PLCJorge Perez CorreaPas encore d'évaluation
- Exposicion Grupo 3Document37 pagesExposicion Grupo 3deniscaza312Pas encore d'évaluation
- Actividad 5 Laboratorios Farmaceuticos Modulo 2Document3 pagesActividad 5 Laboratorios Farmaceuticos Modulo 2Eugenia UrdanetaPas encore d'évaluation
- Salud 3 Historia de La EnfermedadDocument6 pagesSalud 3 Historia de La EnfermedadCarolina Ojeda RomanPas encore d'évaluation
- Comentario Resuelto de Un Texto HumanísticoDocument3 pagesComentario Resuelto de Un Texto HumanísticoelenafezPas encore d'évaluation
- SUNI - Dir - 2021 - by Logan - Sem19Document38 pagesSUNI - Dir - 2021 - by Logan - Sem19Van Soul RiderPas encore d'évaluation
- PolisacáridosDocument17 pagesPolisacáridosMITSUKI TAISHOPas encore d'évaluation
- AxonometriaDocument11 pagesAxonometriaJonbeker JesusPas encore d'évaluation
- Tales de Mileto y Anaxágoras DiapositivasDocument4 pagesTales de Mileto y Anaxágoras DiapositivasJavier JrPas encore d'évaluation
- Ejemplo URDDocument10 pagesEjemplo URDric4rd03098Pas encore d'évaluation
- Informe Previo #3Document10 pagesInforme Previo #3Elian Esteban Chuquillanqui HuamanPas encore d'évaluation
- Ejercicios para Practicar para El ParcialDocument2 pagesEjercicios para Practicar para El ParcialGabriela Alejandra LunaPas encore d'évaluation
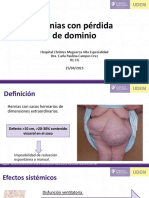
- Hernias Con Pérdida de DominioDocument42 pagesHernias Con Pérdida de DominioCarla CamposPas encore d'évaluation
- El EconomistaDocument12 pagesEl EconomistaMarlon Ben Hur Carlos SantosPas encore d'évaluation
- Parcial 1 HistologiaDocument23 pagesParcial 1 Histologiajuan esteban pinedaPas encore d'évaluation
- Alas para Vivir, Richard BachDocument164 pagesAlas para Vivir, Richard BachZulia PinedaPas encore d'évaluation
- Manual Del XLSTATDocument16 pagesManual Del XLSTATJaime Jose Arias Congrains100% (3)
- PRE UNIVERSITARIO 2021 - Simulacro Semestral Acem 08-08-2021 Bloque CDocument26 pagesPRE UNIVERSITARIO 2021 - Simulacro Semestral Acem 08-08-2021 Bloque CELMER TIGRE HUAMANPas encore d'évaluation
- ExtintoresDocument146 pagesExtintoresMarcello Ross100% (1)
- 1materiales Ceramicos AvanzadosDocument10 pages1materiales Ceramicos AvanzadosJavier LopezPas encore d'évaluation
- Evaluacion Diagnostioca Seguridad e HigieneDocument3 pagesEvaluacion Diagnostioca Seguridad e HigieneBRISA YAMILETH COLIN JIMENEZPas encore d'évaluation
- AmputacionesDocument14 pagesAmputacionesMiguel DiazPas encore d'évaluation