Vous aimerez peut-être aussi
- The Male Reproductive SystemDocument76 pagesThe Male Reproductive SystemteklayPas encore d'évaluation
- Reproductive Systems A. B. C. DDocument6 pagesReproductive Systems A. B. C. DJamie BagundolPas encore d'évaluation
- Zoology Lecture 5Document7 pagesZoology Lecture 5Stephen OcheaPas encore d'évaluation
- Embryology of The Female Reproductive SystemDocument5 pagesEmbryology of The Female Reproductive SystemAndrew LukmanPas encore d'évaluation
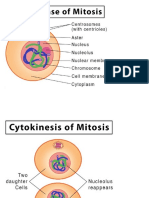
- The Cell CycleDocument9 pagesThe Cell Cyclesalahuddin_md5935Pas encore d'évaluation
- Bone MarrowDocument4 pagesBone MarrowSamra AwanPas encore d'évaluation
- CELL CYCLE + (General Biology)Document17 pagesCELL CYCLE + (General Biology)Jona RodicaPas encore d'évaluation
- BiologyDocument17 pagesBiologyMuskan ShreePas encore d'évaluation
- Chapter 20: The Cardiovascular System: Blood: Embryology Atlas HematopoiesisDocument8 pagesChapter 20: The Cardiovascular System: Blood: Embryology Atlas Hematopoiesisfjestefania6892Pas encore d'évaluation
- Biosciweek6 11Document43 pagesBiosciweek6 11Jesalyn AbrinicaPas encore d'évaluation
- Hema TransesDocument25 pagesHema TransesNikoh Anthony EwayanPas encore d'évaluation
- Bio 001 Cell DivisionDocument8 pagesBio 001 Cell DivisionKnick KnackPas encore d'évaluation
- Reproductive System NotesDocument12 pagesReproductive System NotesTanaya PujarePas encore d'évaluation
- Mitochondria Has A Double Wall Membranous StructureDocument8 pagesMitochondria Has A Double Wall Membranous StructureLalitha BalajiPas encore d'évaluation
- The Male Genital SystemDocument20 pagesThe Male Genital SystemLittleChenaPas encore d'évaluation
- Week 1 - Study QuestionsDocument13 pagesWeek 1 - Study Questionsjess waldenPas encore d'évaluation
- Reduction DivisionaaaaaaDocument2 pagesReduction DivisionaaaaaakimuyyyyPas encore d'évaluation
- Gastrulation and Segmentation ofDocument20 pagesGastrulation and Segmentation ofAina AdesolaPas encore d'évaluation
- Reproductive SystemDocument14 pagesReproductive SystemMichael NyaongoPas encore d'évaluation
- 11 MaleDocument14 pages11 MaleJasmine KaurPas encore d'évaluation
- Cell CycleDocument111 pagesCell Cyclepreyaaa pascuaPas encore d'évaluation
- Emt318 Cytology Note3 2021Document10 pagesEmt318 Cytology Note3 2021Ehigie promisePas encore d'évaluation
- Cell CycleDocument6 pagesCell Cyclewebpixel servicesPas encore d'évaluation
- MeiosisDocument8 pagesMeiosisJohn Chiyu Garde-Labite Azuki100% (1)
- Cell DivisionDocument118 pagesCell DivisionNora AfidaPas encore d'évaluation
- Batterjee Medical College (BMC) FAST (Preparatory Year) Biology Department 2012-2013Document14 pagesBatterjee Medical College (BMC) FAST (Preparatory Year) Biology Department 2012-2013edain84Pas encore d'évaluation
- The ChromosomeHandout - sci8.Q4.LESSON2Document8 pagesThe ChromosomeHandout - sci8.Q4.LESSON2Russ CastilloPas encore d'évaluation
- What Is Meiosis?Document5 pagesWhat Is Meiosis?James DazPas encore d'évaluation
- GametogenesisDocument3 pagesGametogenesisWai KikiPas encore d'évaluation
- Module 2 Meiosis and MitosisDocument88 pagesModule 2 Meiosis and MitosisSevdred CadelinaPas encore d'évaluation
- Cellular Reproduction and GeneticsDocument47 pagesCellular Reproduction and GeneticsJean Marie Macadaeg OrdinarioPas encore d'évaluation
- Embryology GastrulationDocument9 pagesEmbryology GastrulationRivera Gómez América LucíaPas encore d'évaluation
- HAPP Supplementary Notes - Reproductive SystemDocument15 pagesHAPP Supplementary Notes - Reproductive SystemZuhri PorzaPas encore d'évaluation
- Biotechnology NotesDocument21 pagesBiotechnology NotesRommel BauzaPas encore d'évaluation
- Male Histology Trans-Batch 2023Document11 pagesMale Histology Trans-Batch 2023Hanako Sasaki AranillaPas encore d'évaluation
- Mitosis Is A Process Where A Single Cell Divides Into Two Identical Daughter Cells (Cell Division)Document10 pagesMitosis Is A Process Where A Single Cell Divides Into Two Identical Daughter Cells (Cell Division)Holly Mc Kenzie Layson DalmanPas encore d'évaluation
- 2.1.6 Biology Ocr ADocument7 pages2.1.6 Biology Ocr AFatima AfifiPas encore d'évaluation
- Cell CycleDocument19 pagesCell CycleMoffat KaprezzoPas encore d'évaluation
- Cell DivisionDocument2 pagesCell DivisionedukahonPas encore d'évaluation
- Embryology: DR - Kumar Satish Ravi Assist. Prof. & HeadDocument45 pagesEmbryology: DR - Kumar Satish Ravi Assist. Prof. & HeadDr.Kumar Satish RaviPas encore d'évaluation
- SZL B206 Principles of GeneticsDocument28 pagesSZL B206 Principles of Geneticsawuors249Pas encore d'évaluation
- Weeks 3 - 8Document32 pagesWeeks 3 - 8Stefan HutsonPas encore d'évaluation
- MitosisDocument3 pagesMitosisLea Mari JavierPas encore d'évaluation
- Spermatogenesis Is The Process by Which: Seminiferous Tubule With Maturing Sperm.Document2 pagesSpermatogenesis Is The Process by Which: Seminiferous Tubule With Maturing Sperm.Garcia AzirPas encore d'évaluation
- Development of Cardiovascular System PDFDocument58 pagesDevelopment of Cardiovascular System PDFVizhiPas encore d'évaluation
- What Is Mitosis & MeiosisDocument7 pagesWhat Is Mitosis & MeiosisMuhammad VaizPas encore d'évaluation
- InterphaseDocument9 pagesInterphaseJeazel MosendoPas encore d'évaluation
- Organ Reproduksi Laki-Laki: (Organa Genitalia Masculina)Document32 pagesOrgan Reproduksi Laki-Laki: (Organa Genitalia Masculina)Timotius Gatma Buntori PurbaPas encore d'évaluation
- Tdn°2 103019Document9 pagesTdn°2 103019selmi bouzidPas encore d'évaluation
- Meiosis.-Meiosis (del griego μείωσις meíōsis 'disminución')Document6 pagesMeiosis.-Meiosis (del griego μείωσις meíōsis 'disminución')julioPas encore d'évaluation
- Cell Cycle and MitosisDocument49 pagesCell Cycle and Mitosishazharomar958Pas encore d'évaluation
- Male ReproductivehistoDocument9 pagesMale ReproductivehistoaikaPas encore d'évaluation
- Mitosis & Meiosis NotesDocument6 pagesMitosis & Meiosis NotesChris_Barber09100% (1)
- Penugasan Dr. Agung Dewanto, SpOG (K) - Azrul RoeDocument4 pagesPenugasan Dr. Agung Dewanto, SpOG (K) - Azrul RoeAzrul MDPas encore d'évaluation
- Readings Ch04Document11 pagesReadings Ch04Adrian GuzmanPas encore d'évaluation
- Lab Ex1-2 and Chick OrganogenesisiDocument20 pagesLab Ex1-2 and Chick OrganogenesisiLardel CarayPas encore d'évaluation
- 4.2 Cell As A Unit of LifeDocument7 pages4.2 Cell As A Unit of Lifehola adiosPas encore d'évaluation
- Lecture Outline: See Separate Powerpoint Slides For All Figures and Tables Pre-Inserted Into Powerpoint Without NotesDocument58 pagesLecture Outline: See Separate Powerpoint Slides For All Figures and Tables Pre-Inserted Into Powerpoint Without NotesJharaPas encore d'évaluation
- Spermatogenesis 1Document28 pagesSpermatogenesis 1LellllPas encore d'évaluation
- Anatomy and Physiology of Male Reproductive SystemDocument8 pagesAnatomy and Physiology of Male Reproductive SystemAdor AbuanPas encore d'évaluation
- Anaphy Reviewer FINALSDocument4 pagesAnaphy Reviewer FINALSim. EliasPas encore d'évaluation
- CLASS 12 BIO PROJECT SpermatogenesisDocument19 pagesCLASS 12 BIO PROJECT Spermatogenesisanon_38352456389% (9)
- Male Reproductive Physiology - UpToDate - 2020Document18 pagesMale Reproductive Physiology - UpToDate - 2020Karaca AzizPas encore d'évaluation
- Sexual Reproduction in Humans Notes (CAPE)Document29 pagesSexual Reproduction in Humans Notes (CAPE)Desmond JonesPas encore d'évaluation
- Reproductive SystemDocument219 pagesReproductive Systemtzushka_vipPas encore d'évaluation
- OgenisisDocument52 pagesOgenisisBharat ThapaPas encore d'évaluation
- IB Biology Assessment Statements - DrawDocument33 pagesIB Biology Assessment Statements - DrawSchuyler Huff100% (1)
- Histology 18 Male Reproductive SystemDocument49 pagesHistology 18 Male Reproductive SystemAbdul RahmanPas encore d'évaluation
- Intro To Embryology PDFDocument9 pagesIntro To Embryology PDFAdvin BurkePas encore d'évaluation
- Csir Unit 5 Updated PDFDocument10 pagesCsir Unit 5 Updated PDFAmaldev S K100% (2)
- Bio KVS PaperDocument16 pagesBio KVS PaperSanjanaPas encore d'évaluation
- Spermatogenesis, OogenesisDocument16 pagesSpermatogenesis, Oogenesisannita100% (1)
- IB Biology Higher Level Human and Health Physiology NotesDocument41 pagesIB Biology Higher Level Human and Health Physiology NoteshunarsandhuPas encore d'évaluation
- Male and Female Genital Systems 2Document30 pagesMale and Female Genital Systems 2negmm2226Pas encore d'évaluation
- I.J. Singh, Professor Department of Fishery Biology College of Fisheries, G.B.P.U.A.&T., PantnagarDocument39 pagesI.J. Singh, Professor Department of Fishery Biology College of Fisheries, G.B.P.U.A.&T., Pantnagarjoshigauta100% (1)
- Evaluation of Aphrodisiac Activity of Buchanania Axillaris (Linn.) LeavesDocument8 pagesEvaluation of Aphrodisiac Activity of Buchanania Axillaris (Linn.) LeavesDr. Ramadevi DevarakondaPas encore d'évaluation
- Biochemistry of SemenDocument280 pagesBiochemistry of SemenCopper Jean100% (1)
- On The Morphology of The Chromosome Group in Brachystola MagnaDocument18 pagesOn The Morphology of The Chromosome Group in Brachystola MagnaOya KaptanoğluPas encore d'évaluation
- MCQS Human Reproduction Class 12Document13 pagesMCQS Human Reproduction Class 12Shaashwati TandonPas encore d'évaluation
- EM2.Gametogenesis (At)Document52 pagesEM2.Gametogenesis (At)MUDIN ABDELLAPas encore d'évaluation
- Sistem Reproduksi Pria IkrDocument141 pagesSistem Reproduksi Pria IkrNidiyamilhaPas encore d'évaluation
- CLASS 12 BIO PROJECT SpermatogenesisDocument19 pagesCLASS 12 BIO PROJECT Spermatogenesisanon_383524563Pas encore d'évaluation
- KM GametogenesisDocument42 pagesKM GametogenesisKALAMYA BRIAN BWAYOPas encore d'évaluation
- Modul Pintas Tingkatan 5 Peperiksaan Percubaan SPM 2018 Skema Jawapan Biologi Kertas 1 4551/1Document39 pagesModul Pintas Tingkatan 5 Peperiksaan Percubaan SPM 2018 Skema Jawapan Biologi Kertas 1 4551/1VentusPas encore d'évaluation
- 4 AnnotationsDocument8 pages4 Annotationsapi-243334002Pas encore d'évaluation
- OBG DR Pranav PDFDocument183 pagesOBG DR Pranav PDFVinu Poojapranavam100% (6)
- Chapter 21 The Male Reproductive SystemDocument6 pagesChapter 21 The Male Reproductive SystemEllä Pabustan100% (1)