Vous aimerez peut-être aussi
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (588)
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- Everall2017 PDFDocument12 pagesEverall2017 PDFEsteban VegaPas encore d'évaluation
- Bird TourismDocument14 pagesBird TourismEsteban VegaPas encore d'évaluation
- 35 OdonataDocument35 pages35 OdonataEsteban VegaPas encore d'évaluation
- CRISPR 2018 Draft Ethical Principles JiankuiDocument3 pagesCRISPR 2018 Draft Ethical Principles JiankuiEsteban VegaPas encore d'évaluation
- Cit - ScienceDocument17 pagesCit - ScienceEsteban VegaPas encore d'évaluation
- BiodiversityR PDFDocument128 pagesBiodiversityR PDFEsteban VegaPas encore d'évaluation
- Little Etal 2006Document12 pagesLittle Etal 2006Esteban VegaPas encore d'évaluation
- Guía Fotográfica de Familias de Macroinvertebrados Acuáticos de Puerto RicoDocument3 pagesGuía Fotográfica de Familias de Macroinvertebrados Acuáticos de Puerto RicoEsteban VegaPas encore d'évaluation
- FòrmulasDocument2 pagesFòrmulasEsteban VegaPas encore d'évaluation
- The Inner Lives of SpongesDocument3 pagesThe Inner Lives of SpongesEsteban VegaPas encore d'évaluation
- BryopsidaDocument24 pagesBryopsidaEsteban VegaPas encore d'évaluation
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5795)
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (895)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (345)
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (400)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (74)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (266)
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1091)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (121)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- DEPARTMENT OF Dental Anatomy, Embryology and Oral Histology: Dr. Diana Prem VMSDC, SalemDocument25 pagesDEPARTMENT OF Dental Anatomy, Embryology and Oral Histology: Dr. Diana Prem VMSDC, SalemDR. H. DIANA J.HENRY CHRISTOPHERPas encore d'évaluation
- Periodontium (2) - Gingiva: Microscopic Features of GingivaDocument8 pagesPeriodontium (2) - Gingiva: Microscopic Features of Gingivaمحمد محمود القحيفPas encore d'évaluation
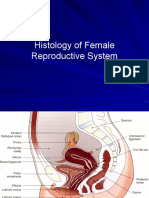
- Female ReproductiveDocument34 pagesFemale ReproductiveAbdallah A. Abu Mattar100% (2)
- Histology of The Gastrointestinal TractDocument10 pagesHistology of The Gastrointestinal TractDwi Rezky AmaliaPas encore d'évaluation
- C - Syiam Amalia Rahmah - Topik 4Document7 pagesC - Syiam Amalia Rahmah - Topik 4Syiam Amalia RahmahPas encore d'évaluation
- Oral Mucous MembraneDocument36 pagesOral Mucous MembraneFourthMolar.com100% (1)
- Surgical Pathology of The Gastrointestinal System Bacterial, Fungal, Viral, and Parasitic Infections PDFDocument234 pagesSurgical Pathology of The Gastrointestinal System Bacterial, Fungal, Viral, and Parasitic Infections PDFBogdan CarabasPas encore d'évaluation
- Medicated Chewing Gum: A Novel Drug Delivery SystemDocument15 pagesMedicated Chewing Gum: A Novel Drug Delivery SystemLiem Cah TunzaPas encore d'évaluation
- Histology: Ust Faculty of Medicine and Surgery Class of 2016Document14 pagesHistology: Ust Faculty of Medicine and Surgery Class of 2016Ashley Beatriz PascualPas encore d'évaluation
- Study Guide 23 - DIGESTIVE SYSTEMDocument33 pagesStudy Guide 23 - DIGESTIVE SYSTEMjed larsen capulong gavinoPas encore d'évaluation
- 234 FarmerDocument6 pages234 FarmerRaghavendra NalatawadPas encore d'évaluation
- (Atlas of Anatomic Pathology) Scott R. Owens, Henry D. Appelman (Auth.) - Atlas of Esophagus and Stomach Pathology-Springer-Verlag New York (2014)Document191 pages(Atlas of Anatomic Pathology) Scott R. Owens, Henry D. Appelman (Auth.) - Atlas of Esophagus and Stomach Pathology-Springer-Verlag New York (2014)Amanda Teaca100% (1)
- CHP 12 - Oral Mucosa-1Document83 pagesCHP 12 - Oral Mucosa-1Areej Fatima100% (2)
- Test Questions: A. Cardiovascular System and LymphaticsDocument24 pagesTest Questions: A. Cardiovascular System and LymphaticsLunna Cruz100% (1)
- Anatomy and Physiology of Gastrointestinal TractDocument23 pagesAnatomy and Physiology of Gastrointestinal TractJasmin Jacob100% (4)
- AJCC Cancer Staging Manual Stomach 7theditionDocument3 pagesAJCC Cancer Staging Manual Stomach 7theditionCamelia CalugariciPas encore d'évaluation
- Advances in Surgical Pathology: Gastric CancerDocument366 pagesAdvances in Surgical Pathology: Gastric CancerBogdan Niculae100% (1)
- Connective TissuesDocument11 pagesConnective TissuesCamille Ann Faigao FamisanPas encore d'évaluation
- Physiology of Mucosae and Mechanism of Transmucosal PermeationDocument48 pagesPhysiology of Mucosae and Mechanism of Transmucosal PermeationArpita Patel100% (1)
- Slide N Label HistologyDocument35 pagesSlide N Label HistologyIsidore Patrick100% (2)
- FinalDocument11 pagesFinalyash rayththaPas encore d'évaluation
- 2019PROCEEDINGBOOKBALIDENCE2019Document855 pages2019PROCEEDINGBOOKBALIDENCE2019Rika LedyPas encore d'évaluation
- Histological and Histochemical Study of The Liver and Gall Bladder of Adult Male Common Carp Cyprinus CarpioDocument5 pagesHistological and Histochemical Study of The Liver and Gall Bladder of Adult Male Common Carp Cyprinus CarpioAbdulkarim Jafar KarimPas encore d'évaluation
- 2000 - GI - Endo InterpretationDocument19 pages2000 - GI - Endo InterpretationambaPas encore d'évaluation
- CP Urinary Bladder Resection 20 4020 PDFDocument15 pagesCP Urinary Bladder Resection 20 4020 PDFBrijeshPas encore d'évaluation
- By: Dr. Susianti, M.SCDocument47 pagesBy: Dr. Susianti, M.SCAnityo NugrohoPas encore d'évaluation
- Atrophic GastritisDocument20 pagesAtrophic GastritisMUJI RIZQIANYPas encore d'évaluation
- Anat 4.3 GIT Histo - ZuluetaDocument8 pagesAnat 4.3 GIT Histo - Zuluetalovelots1234Pas encore d'évaluation
- Histology of Small Intestine For 2nd Year Mbbs (By DR Sundus)Document21 pagesHistology of Small Intestine For 2nd Year Mbbs (By DR Sundus)Mudassar RoomiPas encore d'évaluation
- Obstructive Sleep Aponea SurgeriesDocument638 pagesObstructive Sleep Aponea Surgerieswillyoueverlovemenk100% (2)