Vous aimerez peut-être aussi
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (400)
- Stopping by Woods On A Snowy Evening. FrostDocument1 pageStopping by Woods On A Snowy Evening. Frostnicomaga2000486Pas encore d'évaluation
- A Little While. BronteDocument1 pageA Little While. Brontenicomaga2000486Pas encore d'évaluation
- All Is Well. CloughDocument1 pageAll Is Well. Cloughnicomaga2000486Pas encore d'évaluation
- A Certain Slant of Sunlight. BerriganDocument1 pageA Certain Slant of Sunlight. Berrigannicomaga2000486Pas encore d'évaluation
- Highland Mary. BurnsDocument1 pageHighland Mary. Burnsnicomaga2000486Pas encore d'évaluation
- Belfast Tune. BrodskyDocument1 pageBelfast Tune. Brodskynicomaga2000486Pas encore d'évaluation
- The Alchemist in The City. HopkinsDocument1 pageThe Alchemist in The City. Hopkinsnicomaga2000486Pas encore d'évaluation
- The Frog. BellocDocument1 pageThe Frog. Bellocnicomaga2000486Pas encore d'évaluation
- VVAA. On The Waste LandDocument56 pagesVVAA. On The Waste Landnicomaga2000486Pas encore d'évaluation
- The Figure, Text As Riddle and ForceDocument20 pagesThe Figure, Text As Riddle and Forcenicomaga2000486Pas encore d'évaluation
- On Imagination. WheatleyDocument1 pageOn Imagination. Wheatleynicomaga2000486Pas encore d'évaluation
- Annabel LeeDocument2 pagesAnnabel Leeapi-237662331Pas encore d'évaluation
- "In White": Frost's Early Version of DesignDocument1 page"In White": Frost's Early Version of Designnicomaga2000486Pas encore d'évaluation
- No Man Is An Island. John DonneDocument1 pageNo Man Is An Island. John Donnenicomaga2000486Pas encore d'évaluation
- I Have Loved Hours at Sea. TeasdasleDocument1 pageI Have Loved Hours at Sea. Teasdaslenicomaga2000486Pas encore d'évaluation
- Traveling Through The Dark. StaffordDocument1 pageTraveling Through The Dark. Staffordnicomaga2000486Pas encore d'évaluation
- We Real Cool. BrooksDocument1 pageWe Real Cool. Brooksnicomaga2000486Pas encore d'évaluation
- When I Have Fears. KeatsDocument1 pageWhen I Have Fears. Keatsnicomaga2000486Pas encore d'évaluation
- 70 ArcturusDocument2 pages70 ArcturusnaphelgePas encore d'évaluation
- Richard Cory. Edwin Arlington RobinsonDocument1 pageRichard Cory. Edwin Arlington Robinsonnicomaga2000486Pas encore d'évaluation
- Legend. Hart CraneDocument1 pageLegend. Hart Cranejosesecardi13Pas encore d'évaluation
- Highland Mary. BurnsDocument1 pageHighland Mary. Burnsnicomaga2000486Pas encore d'évaluation
- Booh, FieldDocument1 pageBooh, Fieldnicomaga2000486Pas encore d'évaluation
- It Was Wrong To Do This. Stephan CraneDocument1 pageIt Was Wrong To Do This. Stephan Cranenicomaga2000486Pas encore d'évaluation
- Sonnet 43. Elizabeth Barret BrowningDocument1 pageSonnet 43. Elizabeth Barret Browningnicomaga2000486Pas encore d'évaluation
- A Cloud Withdrew From The Sky. DickinsonDocument1 pageA Cloud Withdrew From The Sky. Dickinsonnicomaga2000486Pas encore d'évaluation
- Personal Intervention Ultima Pantalla - PPT 2011Document1 pagePersonal Intervention Ultima Pantalla - PPT 2011feocorreoPas encore d'évaluation
- 70 ArcturusDocument2 pages70 ArcturusnaphelgePas encore d'évaluation
- A Divine Image. BlakeDocument1 pageA Divine Image. Blakenicomaga2000486Pas encore d'évaluation
- I Have Lived With Shades. HardyDocument2 pagesI Have Lived With Shades. Hardynicomaga2000486Pas encore d'évaluation
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (895)
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (588)
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (74)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (266)
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (344)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (121)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- Super SquatsDocument104 pagesSuper SquatsIt's Just Me100% (3)
- 003 REVIEWER IN SCIENCE 7-SUgarDocument3 pages003 REVIEWER IN SCIENCE 7-SUgarrizza suarezPas encore d'évaluation
- Water PotentialDocument35 pagesWater PotentialLuis GarcíaPas encore d'évaluation
- PPPGGGPPRRDocument18 pagesPPPGGGPPRRRully SetiawanPas encore d'évaluation
- G8 Q4 Bio ReviewedDocument5 pagesG8 Q4 Bio ReviewedSha RonPas encore d'évaluation
- Hcin620 m6 Lab6 Hanifahmutesi-FinalprojectDocument5 pagesHcin620 m6 Lab6 Hanifahmutesi-Finalprojectapi-488096711Pas encore d'évaluation
- Ecosystems Thinking For Mind EcosystemsDocument2 pagesEcosystems Thinking For Mind EcosystemsBala PillaiPas encore d'évaluation
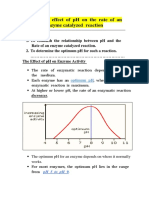
- Effect of PH On Enzyme ActivityDocument12 pagesEffect of PH On Enzyme ActivityAb AbPas encore d'évaluation
- Adhesive Systems in Restorative DentistryDocument6 pagesAdhesive Systems in Restorative DentistryStoian SimonaPas encore d'évaluation
- Oil EOR HandbookDocument24 pagesOil EOR HandbookRicardo Hernan Gomez BPas encore d'évaluation
- Car - Structural Bases For Shaping of DolinesDocument18 pagesCar - Structural Bases For Shaping of DolinesCaegeoPas encore d'évaluation
- Quiz Bowl Competition 2019: Theme: MetabolizeDocument77 pagesQuiz Bowl Competition 2019: Theme: MetabolizeKazuya SenseiPas encore d'évaluation
- Characteristics of A Community PDFDocument15 pagesCharacteristics of A Community PDFAlijanPas encore d'évaluation
- General AnatomiDocument28 pagesGeneral Anatomid25Pas encore d'évaluation
- Induction Presentation Biology A Level and Gcse.204719226Document25 pagesInduction Presentation Biology A Level and Gcse.204719226Cut TirayaPas encore d'évaluation
- PGA102 Presentation 1 (Eng. Ver.)Document18 pagesPGA102 Presentation 1 (Eng. Ver.)Lee Wai KeatPas encore d'évaluation
- Fracturas de ClaviculaDocument11 pagesFracturas de ClaviculaJalil CardenasPas encore d'évaluation
- Northern Blot Analysis For Detection of RNA in PanDocument8 pagesNorthern Blot Analysis For Detection of RNA in PanFati ZoraPas encore d'évaluation
- Domains and Kingdoms: Study GuideDocument6 pagesDomains and Kingdoms: Study GuideosamaPas encore d'évaluation
- Proceedings Biodiversity Research SymposiumDocument156 pagesProceedings Biodiversity Research Symposiumdsn9583Pas encore d'évaluation
- 2019 Introduction To PharmacologyDocument46 pages2019 Introduction To PharmacologyJohn ParajoPas encore d'évaluation
- Bruce Logan MFCDocument9 pagesBruce Logan MFCdavid_tomy_1Pas encore d'évaluation
- 24 5 2022 Ac01.0003860148 Csp-Ics-21254 91-9962016683Document8 pages24 5 2022 Ac01.0003860148 Csp-Ics-21254 91-9962016683Manikandan Ayyasamy100% (1)
- Effect of Square Stepping Exercise Versus Swiss Ball Exercise On Balance in Institutionalized Elderly PopulationDocument6 pagesEffect of Square Stepping Exercise Versus Swiss Ball Exercise On Balance in Institutionalized Elderly Populationastrinila fauziPas encore d'évaluation
- MFK-Drug Management CycleDocument32 pagesMFK-Drug Management CycleLhynda Priarti Latif100% (1)
- Slide CultureDocument2 pagesSlide CultureSoni VishwakarmaPas encore d'évaluation
- Biology Form 4: Chapter 1 (Introduction To Biology)Document2 pagesBiology Form 4: Chapter 1 (Introduction To Biology)Gerard Selvaraj92% (12)
- BRO Fluorescence SpectrosDocument24 pagesBRO Fluorescence SpectrosWaleed El-azabPas encore d'évaluation
- Energy Pyramid - Activity StephanieDocument4 pagesEnergy Pyramid - Activity Stephanieapi-473892313Pas encore d'évaluation
- Pathophysiology of Primary Cataract: Saint Paul University Dumaguete College of Nursing S.Y. 2021-2022Document3 pagesPathophysiology of Primary Cataract: Saint Paul University Dumaguete College of Nursing S.Y. 2021-2022zoie ziazzettePas encore d'évaluation