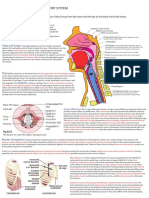
Vous aimerez peut-être aussi
- Medications Used For Smoking CessationDocument12 pagesMedications Used For Smoking Cessationapi-549189480Pas encore d'évaluation
- Electronic Cigarettes: All you need to know about E-Cigarettes and if you should switchD'EverandElectronic Cigarettes: All you need to know about E-Cigarettes and if you should switchPas encore d'évaluation
- Tackling The Smoking Epidemic: IpcrgDocument52 pagesTackling The Smoking Epidemic: IpcrgGoogleyahoointel QuestquestPas encore d'évaluation
- The PERT Concept Q1 A Step-By-Step Approach To Managing Pulmonary EmbolismDocument9 pagesThe PERT Concept Q1 A Step-By-Step Approach To Managing Pulmonary EmbolismntnquynhproPas encore d'évaluation
- APRVDocument20 pagesAPRVivanrom100% (1)
- Pressure Control VentilationDocument17 pagesPressure Control VentilationVijay GadagiPas encore d'évaluation
- Lactic Acidosis: An Update: Jansen Seheult, Gerard Fitzpatrick and Gerard BoranDocument12 pagesLactic Acidosis: An Update: Jansen Seheult, Gerard Fitzpatrick and Gerard Borangasman2003Pas encore d'évaluation
- Pleural Disease LIGTHDocument0 pagePleural Disease LIGTHKaisun TeoPas encore d'évaluation
- Physiology of Mechanical VentilationDocument12 pagesPhysiology of Mechanical VentilationCarlos ZepedaPas encore d'évaluation
- 0221 Community-Acquired-PneumoniaDocument29 pages0221 Community-Acquired-PneumoniaDiego YanezPas encore d'évaluation
- Risk Assessment BiogasDocument6 pagesRisk Assessment BiogasShien LooPas encore d'évaluation
- Pulmonary EmbolismDocument23 pagesPulmonary Embolismaksinu3742Pas encore d'évaluation
- Understanding Care Bundles.9Document4 pagesUnderstanding Care Bundles.9Dianne Erika MeguinesPas encore d'évaluation
- Respiratory Notes (Chris Andersen, ICUPrimaryPrep - Com)Document14 pagesRespiratory Notes (Chris Andersen, ICUPrimaryPrep - Com)PkernPas encore d'évaluation
- Air Pollution Teaching ToolkitDocument74 pagesAir Pollution Teaching ToolkitSuparna Shirke KansakarPas encore d'évaluation
- Case 1Document15 pagesCase 1Hashim QaitoonPas encore d'évaluation
- Pulmonary Function Tests and Lung ResectionDocument9 pagesPulmonary Function Tests and Lung ResectionPrabhakar KumarPas encore d'évaluation
- Mechanical Ventilation GraphicsDocument144 pagesMechanical Ventilation GraphicsSaddamPas encore d'évaluation
- Chest Tube Drainage of The Pleural SpaceDocument10 pagesChest Tube Drainage of The Pleural SpaceFernandaPas encore d'évaluation
- Lung Mechanics and Ventilation-Lecture NotesDocument16 pagesLung Mechanics and Ventilation-Lecture NotesCarlos Eduardo LinaresPas encore d'évaluation
- Bronchopleural Fistula 1frankDocument105 pagesBronchopleural Fistula 1frankroshjamesPas encore d'évaluation
- O2 TerapiDocument36 pagesO2 Terapinurulanisa0703Pas encore d'évaluation
- Clinical Examination of Respiratory SystemDocument37 pagesClinical Examination of Respiratory SystemDipienshu BhatPas encore d'évaluation
- Pulmonary Rehabilitation PPDocument19 pagesPulmonary Rehabilitation PPnashqonashPas encore d'évaluation
- PFT Interpretation DR Giulio DominelliDocument78 pagesPFT Interpretation DR Giulio DominelliFelix ManyerukePas encore d'évaluation
- The Spectrum of TBDocument174 pagesThe Spectrum of TBcmolina84Pas encore d'évaluation
- A Simplified Treatment-Based Approach To TEG and ROTEMDocument7 pagesA Simplified Treatment-Based Approach To TEG and ROTEMCraigbud99Pas encore d'évaluation
- Copd and Equal Pressure PointDocument3 pagesCopd and Equal Pressure Pointleh.mo9315Pas encore d'évaluation
- Gas Transport PDFDocument19 pagesGas Transport PDFMarilia BonorinoPas encore d'évaluation
- Ards Pocket Card 4.7 QRDocument1 pageArds Pocket Card 4.7 QRGenghis SupnetPas encore d'évaluation
- Ventilator Waveforms and The Physiology of Pressure Support PDFDocument21 pagesVentilator Waveforms and The Physiology of Pressure Support PDFRoberto RamirezPas encore d'évaluation
- Cardiovascular SystemDocument40 pagesCardiovascular SystemDouglas Jacques100% (1)
- Portable Oxygen Cylinders: Training and Safety GuidelinesDocument4 pagesPortable Oxygen Cylinders: Training and Safety GuidelinesraoPas encore d'évaluation
- Anatomy of Chest PDFDocument61 pagesAnatomy of Chest PDFRizwan QureshiPas encore d'évaluation
- Lecture Notes On Respiratory Physiology PDFDocument33 pagesLecture Notes On Respiratory Physiology PDFMiles HuiPas encore d'évaluation
- Cardiac Examination Inspection, Palpation & Percussion : Dr. Rajesh Bhat UDocument38 pagesCardiac Examination Inspection, Palpation & Percussion : Dr. Rajesh Bhat URanjith RavellaPas encore d'évaluation
- Pulmonary Function TestingDocument35 pagesPulmonary Function TestingMartha ChasePas encore d'évaluation
- NIPPVDocument35 pagesNIPPVAnusha VerghesePas encore d'évaluation
- The Thromboelastogram (TEG®) : A Five-Minute Primer For The Emergency PhysicianDocument11 pagesThe Thromboelastogram (TEG®) : A Five-Minute Primer For The Emergency PhysicianMarcelliaPas encore d'évaluation
- CRT Exam Review Guide Chapter 5Document9 pagesCRT Exam Review Guide Chapter 5Dharlyn MungcalPas encore d'évaluation
- Clinical Tuberculosis 4th Ed PDFDocument574 pagesClinical Tuberculosis 4th Ed PDFRamonatalePas encore d'évaluation
- An Extension of Barker's Method For Reduction of VLE DataDocument9 pagesAn Extension of Barker's Method For Reduction of VLE Datamurdanetap957Pas encore d'évaluation
- Gas Technology-4th Stage Lecture 2 12 November 2017Document7 pagesGas Technology-4th Stage Lecture 2 12 November 2017muhammadPas encore d'évaluation
- 06 Atkins Chap06Document16 pages06 Atkins Chap06tatianarafaPas encore d'évaluation
- SPE 16984 PA - PDF Performance Predictions For Gas Condensate ReservoirsDocument9 pagesSPE 16984 PA - PDF Performance Predictions For Gas Condensate Reservoirsswaala4realPas encore d'évaluation
- PR 11 TRKDocument20 pagesPR 11 TRKGalihmery DamaiantiPas encore d'évaluation
- Two Phase Flow Safety ValveDocument7 pagesTwo Phase Flow Safety Valversingle81Pas encore d'évaluation
- Glycerin-Water Viscosity Paper - Sent To Ind Eng Chem ResDocument18 pagesGlycerin-Water Viscosity Paper - Sent To Ind Eng Chem ResSulthoni Mukhlis KurniawanPas encore d'évaluation
- Solubility of Carbon Dioxide in Pentadecane, Hexadecane, and Pentadecane + HexadecaneDocument3 pagesSolubility of Carbon Dioxide in Pentadecane, Hexadecane, and Pentadecane + Hexadecanerezagholami87Pas encore d'évaluation
- Modeling of A DC Plasma Torch in Laminar and Turbulent FlowDocument24 pagesModeling of A DC Plasma Torch in Laminar and Turbulent FlowVerayoot1357Pas encore d'évaluation
- Stagnation Pressure Effect On The Supersonic Ow Parameters With Application For Air in NozzlesDocument42 pagesStagnation Pressure Effect On The Supersonic Ow Parameters With Application For Air in NozzlesRou MàissàPas encore d'évaluation
- Saturation Pressure FoysDocument6 pagesSaturation Pressure FoysKalson UmpuPas encore d'évaluation
- Problem Set 1 Data Regression and Numerical Integration - 2013Document3 pagesProblem Set 1 Data Regression and Numerical Integration - 2013April TanjayPas encore d'évaluation
- Equation of StateDocument6 pagesEquation of StateJanardhan CnPas encore d'évaluation
- M25 GpsaDocument24 pagesM25 GpsaPawan ChaturvediPas encore d'évaluation
- Isentropic Expansion of Condensing SteamDocument9 pagesIsentropic Expansion of Condensing SteamHoustonAbducteePas encore d'évaluation
- Glycine Proton Transfer Enthalpy Summary and Data Analysis GuideDocument4 pagesGlycine Proton Transfer Enthalpy Summary and Data Analysis GuideRuby RichiezPas encore d'évaluation
- Calculation of Phase Envelopes and Critical Points For Multicomponent MixturesDocument10 pagesCalculation of Phase Envelopes and Critical Points For Multicomponent Mixturesflavio_cordero_1Pas encore d'évaluation
- Polymer Engineering ExerciseDocument10 pagesPolymer Engineering ExerciseMiguel A. GraneroPas encore d'évaluation
- Sonnenschein 1986 5Document3 pagesSonnenschein 1986 5niko_sonnenscheinPas encore d'évaluation
- Rodaks Hematology, 5EDocument917 pagesRodaks Hematology, 5Emr microsdgamer je100% (2)
- Digital Edition July 2020Document1 pageDigital Edition July 2020akbianchessiPas encore d'évaluation
- Genetics Lands in Primary Care Inboxes: ME Multiplex Panel: Debating The TradeoffsDocument74 pagesGenetics Lands in Primary Care Inboxes: ME Multiplex Panel: Debating The TradeoffsakbianchessiPas encore d'évaluation
- Sarampo ImunossupressaoDocument15 pagesSarampo ImunossupressaoakbianchessiPas encore d'évaluation
- Game of Clones - How Measles Remodels The B Cell Landscape Sarampo VacinacaoDocument4 pagesGame of Clones - How Measles Remodels The B Cell Landscape Sarampo VacinacaoakbianchessiPas encore d'évaluation
- Cap 2019 12 01Document77 pagesCap 2019 12 01akbianchessiPas encore d'évaluation
- HbA1c Platforms Studied For Lipemia InterferenceDocument7 pagesHbA1c Platforms Studied For Lipemia InterferenceakbianchessiPas encore d'évaluation
- New DNA Test Quickly Identifies Multidrug - TranslatedDocument4 pagesNew DNA Test Quickly Identifies Multidrug - TranslatedakbianchessiPas encore d'évaluation
- Genetics Lands in Primary Care Inboxes: ME Multiplex Panel: Debating The TradeoffsDocument74 pagesGenetics Lands in Primary Care Inboxes: ME Multiplex Panel: Debating The TradeoffsakbianchessiPas encore d'évaluation
- Blood How CollDocument5 pagesBlood How Collakbianchessi100% (1)
- Absolute Band Counts in Febrile Infants - Know Your Laboratary (PediatricsDocument1 pageAbsolute Band Counts in Febrile Infants - Know Your Laboratary (PediatricsakbianchessiPas encore d'évaluation
- LMI Cofator de Ristocetina BioFlash JornalDocument2 pagesLMI Cofator de Ristocetina BioFlash JornalakbianchessiPas encore d'évaluation
- Fever in Infants and ChildrenDocument24 pagesFever in Infants and ChildrenakbianchessiPas encore d'évaluation
- Effect of Temperature On The P50Value For Human BloodDocument5 pagesEffect of Temperature On The P50Value For Human BloodakbianchessiPas encore d'évaluation
- Preanalytical Errors Introduced by Sample-Transportation Systems - A Means To Asses ThemDocument2 pagesPreanalytical Errors Introduced by Sample-Transportation Systems - A Means To Asses ThemakbianchessiPas encore d'évaluation
- CAP Artigo CoagulacaoDocument1 pageCAP Artigo CoagulacaoakbianchessiPas encore d'évaluation
- Estimation of The Minimal Preanalytical Uncertainty For 15 Clinical Chemistry Serum AnalytesDocument7 pagesEstimation of The Minimal Preanalytical Uncertainty For 15 Clinical Chemistry Serum AnalytesakbianchessiPas encore d'évaluation
- EX Cleft SentencesDocument4 pagesEX Cleft SentencesakbianchessiPas encore d'évaluation
- Diabete Ndfs 2011 CDCDocument12 pagesDiabete Ndfs 2011 CDCakbianchessiPas encore d'évaluation
- Discordance Between Measured and Calculated Total Carbon DioxideDocument4 pagesDiscordance Between Measured and Calculated Total Carbon DioxideakbianchessiPas encore d'évaluation
- Essential Guide To FDADocument33 pagesEssential Guide To FDAakbianchessiPas encore d'évaluation
- Diabete Ndfs 2011 CDCDocument12 pagesDiabete Ndfs 2011 CDCakbianchessiPas encore d'évaluation
- Pentra XL80Pentra BrochDocument8 pagesPentra XL80Pentra BrochakbianchessiPas encore d'évaluation
- Current Analytical Approaches To Measuring Blood AnalytesDocument5 pagesCurrent Analytical Approaches To Measuring Blood AnalytesakbianchessiPas encore d'évaluation
- Pentra 60 OTDocument2 pagesPentra 60 OTakbianchessiPas encore d'évaluation
- Pentra XL80Pentra BrochDocument8 pagesPentra XL80Pentra BrochakbianchessiPas encore d'évaluation
- Pentra XL80Pentra BrochDocument8 pagesPentra XL80Pentra BrochakbianchessiPas encore d'évaluation
- MAGEBA Elastomeric BearingsDocument18 pagesMAGEBA Elastomeric BearingsJose ManzanarezPas encore d'évaluation
- Physical Science (Specialization) Reviewer 494 Items With Rationalization PDFDocument118 pagesPhysical Science (Specialization) Reviewer 494 Items With Rationalization PDFKaren DellatanPas encore d'évaluation
- Stainless Steel in Fire Final Summary Report March 08Document121 pagesStainless Steel in Fire Final Summary Report March 08twinpixtwinpixPas encore d'évaluation
- Shaft Model With Mathlook EquationsDocument112 pagesShaft Model With Mathlook EquationsMoisés MachadoPas encore d'évaluation
- Durethan B30S 000000: PA 6, Non-Reinforced, Injection Molding ISO Shortname: ISO 1874-PA 6, GR, 14-030Document3 pagesDurethan B30S 000000: PA 6, Non-Reinforced, Injection Molding ISO Shortname: ISO 1874-PA 6, GR, 14-030zoranPas encore d'évaluation
- Sam Townsend CV 2011Document2 pagesSam Townsend CV 2011Sam TownsendPas encore d'évaluation
- E2906E2906M-13 Standard Practice For Acoustic Pulse Reflectometry Examination of Tube BundlesDocument6 pagesE2906E2906M-13 Standard Practice For Acoustic Pulse Reflectometry Examination of Tube BundlesMohamed100% (1)
- (CO5) Chemical EquilibriumDocument35 pages(CO5) Chemical EquilibriumAya Evangelista AlmandresPas encore d'évaluation
- Aggregates in ConcreteDocument16 pagesAggregates in Concretevinaykrishna123Pas encore d'évaluation
- Flame TestDocument9 pagesFlame TestChristopher YepmoPas encore d'évaluation
- 3130507-Group Wise Topics For Online PresentationDocument2 pages3130507-Group Wise Topics For Online PresentationKaushal BaldhaPas encore d'évaluation
- 03 Uppercat Free Upcat KeyDocument40 pages03 Uppercat Free Upcat Keyadrian suppPas encore d'évaluation
- EHB en File 10.5.1 PressureTemperature Ratings Acc To JIS B 2220 2239Document4 pagesEHB en File 10.5.1 PressureTemperature Ratings Acc To JIS B 2220 2239wholenumberPas encore d'évaluation
- The Epoxy BookDocument37 pagesThe Epoxy BookEvTech PhilPas encore d'évaluation
- C26x Enm C57a PDFDocument1 284 pagesC26x Enm C57a PDFmn090078dPas encore d'évaluation
- EPSGCODES-Authority Codes For Coordinate Reference SystemsDocument249 pagesEPSGCODES-Authority Codes For Coordinate Reference SystemsmizonexPas encore d'évaluation
- Is - 00191 - 2007 PDFDocument14 pagesIs - 00191 - 2007 PDFSangita GhaisasPas encore d'évaluation
- Natural Philosophy: Language Watch EditDocument7 pagesNatural Philosophy: Language Watch Editlin ًPas encore d'évaluation
- Conceptual Fluid QuestionsDocument42 pagesConceptual Fluid QuestionsAmr Faisal RaghebPas encore d'évaluation
- Us7201104 PDFDocument7 pagesUs7201104 PDFThamuze UlfrssonPas encore d'évaluation
- AME101 LectureNotesDocument142 pagesAME101 LectureNotesNagarjunavarma GannaPas encore d'évaluation
- Mil STD 1541aDocument42 pagesMil STD 1541aramadhany_kizunaPas encore d'évaluation
- Blast Resistant Design With Structural Steel PDFDocument6 pagesBlast Resistant Design With Structural Steel PDFMatthieuPas encore d'évaluation
- Com Flor 60 BrochureDocument28 pagesCom Flor 60 BrochureMilena JanicijevicPas encore d'évaluation
- Neutron LogDocument56 pagesNeutron LogYansyah Oktriono Putra67% (3)
- ORMOCERIDocument6 pagesORMOCERIBogdanPas encore d'évaluation
- Number Patterns and Sequences and Decimals HotsDocument3 pagesNumber Patterns and Sequences and Decimals Hotszarina binti jusohPas encore d'évaluation
- GE Lighting Systems HM-1000 Asymmetrical High Mast Series Spec Sheet 12-74Document2 pagesGE Lighting Systems HM-1000 Asymmetrical High Mast Series Spec Sheet 12-74Alan Masters100% (1)
- Number System Question AnswersDocument4 pagesNumber System Question AnswersjaynipsPas encore d'évaluation
- Standard S Ific Ion For: - !' N G :.. F.Ld'.Tita"Ilj.MDocument6 pagesStandard S Ific Ion For: - !' N G :.. F.Ld'.Tita"Ilj.MIgnacio Hiram M RoquePas encore d'évaluation