Académique Documents
Professionnel Documents
Culture Documents
Apostila 1 - 2013
Transféré par
Marcus Vinicius AlvesCopyright
Formats disponibles
Partager ce document
Partager ou intégrer le document
Avez-vous trouvé ce document utile ?
Ce contenu est-il inapproprié ?
Signaler ce documentDroits d'auteur :
Formats disponibles
Apostila 1 - 2013
Transféré par
Marcus Vinicius AlvesDroits d'auteur :
Formats disponibles
Coordenao Geral Comisso Organizadora Andr Bedendo de Souza Marcus Vinicius Costa Alves
Professores Responsveis Profa. Dra. Isabel Marian Hartmann de Quadros Prof. Dr. Jair Ribeiro Chagas
Coordenadores de Mdulo Mdulo Introdutrio Marina Rezende e Guilherme Julian Mdulo de Modelos Experimentais Csar Augusto de Oliveira Coelho Mdulo de Memria Vanessa Manchim Favaro Mdulo de Neuropsicologia Larissa Botelho Gaa Mdulo de Transtornos Psiquitricos Mayra Machado e Luiz Dieckmann Mdulo de Plantas e SNC Joo Vitor Pildervasser Mdulo de Drogas de Abuso Juara Zaparoli Mdulo de Estresse Mariella Bodemeier Loayza Careaga Mdulo de Cronobiologia Bruno Jacson Martynhak Mdulo de Sono Flvia de Mattos Egydio Mdulo de Exerccio Fsico Valdir Aquino
Coordenador da Tutoria Cientfica Gabriel Natan Pires
-2-
Patrocinadores
Apoio
-3-
Prefcio...............................................................................................................................5 Mdulo Introdutrio............................................................................................................6 Neuroanatomia Funcional.....................................................................................6 Neurofisiologia........................................................................................................32 Psicofarmacologia.................................................................................................45 Modelos Experimentais....................................................................................................74
-4-
O Curso de Vero em Psicobiologia uma iniciativa de discentes e docentes do Departamento de Psicobiologia da UNIFESP e que desde o ano de 2001 vem anualmente deixando sua marca entre os profissionais das neurocincias do Brasil. E com essa perspectiva, a de deixar ainda mais estabelecida a marca desta dedicao e iniciativa, que com imenso prazer ns recebemos vocs alunos para a XIII edio do curso. Nosso objetivo fornecer as bases para o pensamento cientfico em Psicobiologia. Desta maneira, o curso pretende apresentar conhecimentos, mtodos experimentais e possibilitar a vivncia acadmica. O Curso de Vero em Psicobiologia busca integrar e expandir o conhecimento cientfico nacionalmente. E foi com a vontade de unir diversos conhecimentos, proporcionando um pensamento amplo sobre o tema que o Departamento de Psicobiologia foi criado em 1971, como um setor do Departamento de Farmacologia da antiga Escola Paulista de Medicina, tornando-se um departamento independente pouco tempo depois (1973). Desde ento vm sendo realizados inmeros projetos de pesquisas de alto impacto na rea da sade. Esperamos que nesta apostila e ao longo do curso vocs encontrem no apenas respostas, mas principalmente, novas perguntas. As bases biolgicas do comportamento abraam uma infinidade de tpicos que podem motivar curiosidade para um cientista em formao, e o nosso curso almeja despertar tal interesse. A apostila com o contedo do curso ser divida em trs partes: I Introduo Neuroanatomia, Psicofarmacologia e Neurofisiologia e Modelos Experimentais; II Memria, Neuropsicologia, Transtornos Psiquitricos e Plantas e SNC; e III Drogas de Abuso, Estresse, Sono e Exerccio Fsico.
Sejam muito bem vindos! Desejamos um bom proveito dessa experincia!
Coordenao do XII Curso de Vero em Psicobiologia Andr Bedendo de Souza, Marcus Vinicius Costa Alves Profa. Dra. Isabel Marian Hartmann de Quadros e Prof. Dr. Jair Ribeiro Chagas
-5-
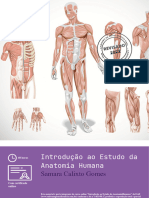
NEUROANATOMIA FUNCIONAL
1. CLASSIFICAO E ORGANIZAO GERAL DO SISTEMA NERVOSO
Com base em critrios anatmicos, o sistema nervoso (SN) humano pode ser classificado em Sistema Nervoso Perifrico (SNP), formado por gnglios, nervos e terminaes nervosas sensitivas e motoras e Sistema Nervoso Central (SNC). O SNC compreende estruturas do SN que esto envoltas por um esqueleto sseo (cavidade craniana e canal vertebral): o encfalo e a medula espinhal. O encfalo, por sua vez, pode ser dividido em seis partes principais: bulbo, ponte e mesencfalo (os quais compem o tronco enceflico), cerebelo, diencfalo e os hemisfrios cerebrais (figura 1). Alm disso, o SNC possui cavidades contendo lquido cefalorraquideano e que formam o sistema ventricular, que composto por quatro ventrculos: os ventrculos laterais esquerdo e direito, presentes no telencfalo; o terceiro ventrculo, localizado no diencfalo; e no tronco cerebral encontramos o quarto ventrculo. Este ltimo continua formando o canal central da medula.
Figura 1: Diviso anatmica do sistema nervoso. SN: sistema nervoso; SNC: sistema nervoso central; SNP: sistema nervoso perifrico (adaptado de Bear MF, 2007).
-6-
Embriologicamente o SN deriva-se do ectoderma, o folheto embrionrio mais externo (folhetos embrionrios so camadas celulares presentes no embrio e que iro diferenciar-se nos diversos rgos). Ao longo do desenvolvimento do embrio, o ectoderma ir sofrer diversas modificaes, principalmente na regio da notocorda, formando a placa neural. A placa neural se torna espessa e origina o sulco neural, que se fechar do meio para as extremidades formando o tubo neural. Este ltimo dilata-se, em sua regio rostral, e d origem s vesculas precursoras do SNC: prosencfalo, mesencfalo e rombencfalo; a regio do tubo neural que no passa por mudanas de calibre (dilatao) d origem medula espinhal. Com o desenvolvimento, as trs vesculas do origem a cinco (telencfalo, diencfalo, mesencfalo, metencfalo e milencfalo), e aparecem trs curvaturas no embrio (ceflica, cervical e pontina), sendo que a ceflica a nica a permanecer at a vida adulta (figura 2). Nos pontos em que o ectoderma que envolve o sulco neural se encontra, formada a crista neural, precursora dos elementos do SNP.
Figura 2: Diviso embriolgica do Sistema Nervoso. A: etapas do desenvolvimento do ectoderma. B: vesculas precursoras do Sistema Nervoso Central. C: curvaturas presentes no embrio (modificado de Machado ABM, 2006).
O desenvolvimento das clulas que compem o SN se d a partir do tubo neural: os neuroblastos, que originam os diferentes tipos de neurnios; os gliobastos, dos quais a macrglia (astrcitos, oligodendrcitos e clulas de Schwann) originada; e as clulas ependimrias, que formam o revestimento interno dos ventrculos cerebrais e plexo coride. De todos esses precursores, apenas as clulas ependimrias no se desenvolvem e permanecem como clulas tronco embrionrias. No SNC adulto existem basicamente duas classes distintas de clulas: os neurnios e as clulas da glia. Embora as clulas gliais sejam cerca de 10 vezes mais numerosas do que as clulas neuronais, as caractersticas funcionais dos neurnios fazem desta classe a mais estudada na
-7-
neurocincia.
1.1. Neurnios: Unidades Funcionais
Estruturalmente, um neurnio tpico apresenta trs regies morfologicamente definidas: o corpo celular, os dendritos e o axnio (figura 3A). A membrana neuronal a camada mais externa do neurnio, a qual separa o meio intracelular do meio extracelular.
Figura 3: Representao esquemtica do neurnio. A: morfologia da clula neuronal. B: classificao dos neurnios em relao ao nmeros de processos que se originam do corpo celular (adaptado de Machado ABM, 2006).
O corpo celular do neurnio contm as mesmas organelas presentes nas demais clulas do organismo, sendo o ncleo (local de armazenamento da informao gentica) a organela principal. Tudo que est contido dentro da membrana no corpo celular, exceto o ncleo, pode ser denominado de citoplasma. Devido presena de material gentico e organelas responsveis pela sntese e degradao de protenas, o corpo celular o centro metablico do neurnio. Uma caracterstica nica aos neurnios a presena de axnio: estrutura altamente especializada que transmite informaes no SN. Estruturalmente apresenta, alm da membrana plasmtica, o axoplasma (citoplasma axnico) no qual encontram-se protenas provenientes do corpo celular (j que no axnio no h a maquinaria necessria para produzi-las). Os axnios variam em comprimento, podendo se estender por mais de um metro. Os ramos de um nico axnio podem formar sinapses com milhares de neurnios, inclusive com o prprio neurnio de
-8-
origem. A regio final do axnio denominada axnio terminal ou boto terminal. Nesta regio o axnio entra em contato com outra clula transmitindo a informao nervosa. De maneira geral, enquanto o axnio o elemento de sada do neurnio, os dendritos so, teoricamente, elementos de entrada. Isto se deve ao fato da membrana dendrtica possuir diversas protenas responsveis pela captao da informao transmitida. Entretanto, sua funo no se restringe a receber informaes apenas, os dendritos podem compartilhar juntamente com os axnios a habilidade de transmitir sinais eltricos. Os dendritos de um nico neurnio so coletivamente denominados rvore dendrtica. Morfologicamente os neurnios podem ser classificados de diversas maneiras: - Pelo nmero de prolongamentos que se originam do corpo celular: o Unipolares: um nico prolongamento que pode funcionar como superfcie receptiva ou terminal liberador, o o Bipolares: possuem dois prolongamentos funcionalmente especializados, Pseudo-unipolares: possuem um prolongamento nico que emerge do corpo celular e se divide em dois, ambos atuando como axnio e; o Multipolares: possuem um axnio e muitos dendritos, estrutura que permite grande nmero de aferncias sinpticas (figura 3B). - Pela estrutura da rvore dentrtica: o Clulas estreladas e clulas piramidais: presentes no crtex cerebral; Neurnios sensoriais, interneurnios e neurnios motores e; Neurnios de projeo (longos axnios) e neurnios de circuito local (curtos axnios). - Baseada em suas conexes: o - De acordo com o comprimeto axonal: o
1.2. Clulas da Glia
O nome glia provm da palavra grega glue, que significa cola, pois essa era a funo atribuda a essa classe de clulas: acreditava-se que a funo da glia era manter os neurnios unidos. Atualmente sabe-se que estas clulas possuem diversas funes importantes no SNC como, por exemplo: - proporcionar sustentao, dando firmeza e estrutura ao crebro, alm de separar e ocasionalmente isolar grupos de neurnios entre si; - formao da bainha de mielina, que um material isolante presente em grande parte dos neurnios, importante para a manuteno da propagao do sinal neuronal; + - tamponamento e manuteno da concentrao dos ons potssio (K ) no meio extracelular;
-9-
- captao e remoo de transmissores qumicos liberados pelos neurnios; - orientao do movimento migratrio neuronal e do crescimento axonal durante o desenvolvimento cerebral; - participao na criao do revestimento dos capilares cerebrais criando a barreira hematoenceflica que impede o acesso de substncias txicas ao crebro; - possvel participao na nutrio de clulas neuronais de vertebrados. So divididas em duas grandes classes: a micrglia e a macrglia. Existem trs subtipos de clulas da macrglia que predominam no sistema nervoso de vertebrados: os oligodendrcitos, as clulas de Schwann e os astrcitos (figura 4).
Figura 4: Representao esquemtica das clulas da glia (adaptado de Kandel, ER, 2000).
1.3. Micrglia
A micrglia, cujas clulas tm origem mesodrmica, o grande mediador da resposta imunolgica no SNC. Consideradas funcionalmente similar aos leuccitos. Contudo, so as clulas menos compreendidas do SNC. No se sabe ao certo qual o papel da micrglia no estado fisiolgico normal. Possivelmente, essas clulas desempenham um papel vital no desenvolvimento do SN e na patognese de doenas do SNC.
1.4. Oligodendrcitos e Clulas de Schwann
A bainha de mielina produzida pelos oligodendrcitos. Cada oligodendrcito capaz de revestir e manter a bainha de mielina em diversos neurnios. A bainha de mielina no reveste
- 10 -
totalmente os axnios, cada segmento mielinizado chamado de internodo, pois ao final de cada segmento existe uma poro no revestida denominada nodo de Ranvier (figura 3A). Fisiologicamente, a mielina possui propriedade de isolamento, dessa forma o potencial de ao que percorre o neurnio capaz de "saltar" de n para n, garantindo uma conduo rpida (conduo saltatria). J no SNP, a mielina produzida pelas clulas de Schwann, sendo que cada uma dessas clulas reveste um nico segmento axonal. O tipo de mielina produzida pelos oligodendrcitos e clulas de Schwann possui como componente principal a Protena Bsica de Mielina (PBM), no entanto, o subtipo presente em ambas difere. A mielina produzida pelos oligodendrcitos possui a PBM do tipo I, o qual impede a regenerao do axnio frente a leses. O mesmo no ocorre na mielina produzida perifericamente.
1.5. Astrcitos
Os astrcitos so as clulas gliais mais numerosas, possuem esse nome pelo corpo celular com formato irregular e rudemente semelhante a uma estrela. Possuem longos processos, sendo que suas terminaes podem fazer contato com neurnios do crebro e medula espinhal, transportando nutrientes necessrios para essas clulas, ou com os vasos sanguneos cerebrais. Nesta ltima, os astrcitos estimulam as clulas endoteliais dos vasos a produzirem uma juno do tipo tight. As junes tight limitam a passagem de substncias do vaso sanguneo para as demais clulas do SNC, e criam uma barreira de proteo entre o sistema vascular e o crebro. Estas clulas tambm emitem projees junto aos axnios, formando um envoltrio ao redor dos nodos de Ranvier e participam tambm do processo de migrao neuronal durante o desenvolvimento e no tamponamento e manuteno das concentraes inicas do meio extracelular, o que pode ser importante na formao, maturao, manuteno e eficcia das sinapses.
2. DIVISO ANATOMO-FUNCIONAL
2.1. Telencfalo
O telencfalo compreende os dois hemisfrios cerebrais, direito e esquerdo, os quais so unidos por uma larga faixa de fibras comissurais: o corpo caloso (maior feixe de fibras do SN que permite a transferncia de informaes de um hemisfrio cerebral para outro).
- 11 -
Cada hemisfrio possui trs plos: frontal, occipital e temporal; e trs faces: spero-lateral (convexa), medial (plana) e inferior ou base do crebro (irregular). Sua superfcie apresenta depresses denominadas sulcos, os quais possibilitam o aumento da superfcie cerebral sem alterar o volume, e que delimitam os giros cerebrais. O padro de sulcos e giros varivel, no entanto alguns apresentam-se de forma constante, recebendo denominaes especiais e ajudando a delimitar os lobos e reas cerebrais. Os dois sulcos mais importantes so o sulco lateral (de Sylvius), que separa os lobos frontal, temporal e parietal, e o sulco central (de Rolando), que separa os giros pr-central (rea relacionada motricidade) e ps-central (rea relacionada sensibilidade geral) (figura 5A).
Figura 5: Anatomia macroscpica do telencfalo. A: principais giros e sulcos. B: representaco da diviso dos lobos (adaptado de Bear MF, 2007).
Os lobos cerebrais recebem denominaes de acordo com os ossos do crnio aos quais se relacionam: lobo frontal, lobo parietal, lobo temporal e lobo occipital. Existe, ainda, um quinto lobo, a nsula, situado profundamente no sulco lateral e sem relao com os ossos cranianos (figura 5B). Embora a diviso dos lobos seja apenas anatmica, exceto pelo lobo occipital e sua relao direta com a viso, os sulcos e giros presentes em cada lobo apresentam caractersticas anato-funcionais. Segue abaixo alguns exemplos de sulcos e giros e suas relaes funcionais: 1) Lobo frontal - giro pr-central: onde localiza-se a rea motora principal do crebro; giro frontal inferior (subdividido em trs partes - orbital, triangular opercular); no hemisfrio esquerdo este giro denominado giro ou rea de Brocca, constituindo-se, na maioria dos indivduos, o centro cortical da palavra falada. 2) Lobo temporal - giro temporal transverso anterior: centro cortical da audio; giro parahipocampal e giro do cngulo que fazem parte do lobo lmbico, relacionado com o comportamento emocional e o controle do sistema nervoso autnomo. 3) Lobo parietal - giro ps-central: rea somestsica (uma das mais importantes reas sensitivas do crtex). 4) Lobo occipital - sulco calcarino: centro cortical da viso.
- 12 -
2.1.1. Crtex Cerebral
O crtex cerebral compreende a camada de substncia cinzenta que reveste o centro branco medular do crebro. Localiza-se na superfcie e apresenta vrias circunvolues, os sulcos e giros meniconados anteriormente, permitem que uma grande rea esteja presente dentro da cavidade craniana. No crtex chegam impulsos provenientes de todas as vias da sensibilidade que se tornam conscientes e so interpretados; dele saem os impulsos nervosos que iniciam e comandam os movimentos voluntrios e com ele esto relacionados os fenmenos psquicos superiores. Filogeneticamente, o crtex cerebral humano pode ser divido em arquicrtex, paleocrtex e neocrtex. O arquicrtex e paleocrtex compreendem estrututras do hipocampo e giro parahipocampal, e o neocrtex o crtex propriamente dito. Estruturalmente, o crtex cerebral pode ser dividido em isocrtex e alocrtex. O isocrtex apresenta seis camadas, distintas pela composio celular presente, e corresponde ao neocrtex; j o alocrtex corresponde ao arqui e paleo crtex e sua citoarquitetura. As seis camadas que compem o crtex (nomeadas de I a VI, partindo da camada mais externa para a mais interna) distinguem-se pelos diferentes tipos neuronais. A camada I, ou molecular, a mais superficial, composta essencialmente por fibras de direo horizontal e neurnios horizontais (clulas de Cajal) de associao intracortical. Na camada II, camada granular externa, contm neurnios granulares, reconhecidos como os principais interneurnios cortical e tambm as clulas receptoras do crtex cerebral. Na camada III, camada piramidal externa, h o predomnio de clulas piramidais. Estas possuem axnios que se direcionam de forma descendente, compondo as fibras eferentes do crtex. A camada IV, camada granular interna, possui caractersticas similares camada II. Na camada V, camada piramidal interna, tambm predomina as clulas piramidais, portanto as camadas III e V apresentam caractersticas essencialmente efetoras. Por fim, a camada VI, camada de clulas fusiformes ou multiformes, composta por neurnios que possuem prolongamentos axonais em direo ao centro branco-medular, tendo caractersticas efetoras. Funcionalmente o crtex pode ser classificado em dois grandes grupos: reas de projeo e reas de associao. As reas de projeo recebem ou do origem a fibras relacionadas diretamente com a sensibilidade ou motricidade, e so consideradas reas primrias. As reas de associao esto relacionadas com funes psquicas complexas, podendo ser divididas em reas secundrias e tercirias: a) reas secundrias (unimodais) ou esto a relacionadas motricidade indiretamente (suas conexes com se uma fazem determinada modalidade sensorial com
predominantemente com a rea primria da mesma funo) e; b) reas tercirias (supramodais) no se ocupam do processamento motor ou sensitivo, mas esto envolvidas com atividades cognitivas superiores (mantm conexes com vrias reas
- 13 -
unimodais e com outras reas supramodais). reas de Projeo: reas Sensitivas Primrias a) rea somestsica ou rea da sensibilidade somtica geral est localizada no giro pscentral e recebe informaes vindas do tlamo a respeito de temperatura, dor, presso, tato e propriocepo consciente da metade oposta do corpo. b) rea visual: localizada nos lbios do sulco calcarino e recebe aferncias do corpo geniculado lateral. c) rea auditiva: localizada no giro temporal transverso anterior e recebe aferncias do corpo geniculado medial. d) rea vestibular: localizada no lobo parietal, e est relacionada com a rea de projeo da sensibilidade proprioceptiva, pois localiza-se prximo a ao territrio da rea somestsica correspondente face. Tambm apresenta papel importante a orientao consciente no espao. e) rea olfatria: em humanos ocupa uma pequena rea situada na parte anterior do ncus e do giro para-hipocampal. f) rea gustativa: localizada na poro inferior do giro ps-central, prximo a nsula, em uma regio adjacente parte da rea somestsica correspondente lngua. Leses nessas reas provocam uma diminuio da gustao na metade oposta da lngua. rea motora primria Ocupa a parte posterior do giro pr-central. Suas principais conexes aferentes so com o tlamo (atravs do qual recebe informaes do cerebelo), com a rea somestsica e com as reas pr-motora e motora suplementar. Em humanos, origina as fibras responsveis pela motricidade voluntria. reas de Associao: reas de associao secundrias sensitivas So trs as reas conhecidas: rea somestsica secundria (lbulo parietal superior), rea visual secundria (lobos temporal e occipital) e rea auditiva secundria (lobo temporal). Recebem informaes das reas primrias correspondentes e repassam informaes recebidas para outras reas do crebro, especialmente s reas supramodais. Leses nessas reas levam a agnosias visuais, auditivas e somestsicas (incapacidade de reconhecer objetos pela viso, som emitido ou tato, embora possam ser reconhecidos por outro sentido que no o afetado) e a afasias (dificuldades de compreenso de sons de linguagem). reas de associao secundria motoras Localizam-se adjacentemente rea motora primria, com a qual se relacionam. Leses
- 14 -
dessas reas frequentemente causam apraxia (incapacidade de executar determinados atos voluntrios, sem que exista qualquer dficit motor). Neste caso, a leso est nas reas corticais de associao relacionadas com o planejamento dos atos voluntrios e no na execuo destes. a) rea motora suplementar: localizada na face medial do giro frontal superior mantm conexes com o corpo estriado, via tlamo, e com a rea motora primria. Funcionalmente, relaciona-se com a concepo ou planejamento de sequncias complexas de movimentos, sendo ativada juntamente com a rea motora primria quando esses movimentos so executados. b) rea pr-motora: localizada frente da rea motora primria tem projees para esta e para a formao reticular, recebendo aferncias do cerebelo, via tlamo, e de vrias reas de associao do crtex. Funcionalmente, coloca o corpo em uma postura preparatria para os movimentos mais delicados. c) rea de Brocca: localizada nas partes opercular e triangular do giro frontal inferior responsvel pela programao da atividade motora relacionada com a expresso da linguagem. Leses dessa rea resultam em afasias (distrbios de linguagem expressiva). reas de associao tercirias (Supramodais) No se relacionam isoladamente com nenhuma modalidade sensorial (da o termo supramodal). Recebem e integram as informaes sensoriais j elaboradas por todas as reas secundrias e so responsveis tambm pela elaborao das diversas estratgias comportamentais. a) rea pr-frontal: compreende a parte anterior no-motora do lobo frontal que recebe fibras de todas as demais reas de associao do crtex, ligando-se ainda ao sistema lmbico. Leses nesta rea alteram as funes cognitivas normais, levando perda de senso de responsabilidades sociais, dificuldade na manuteno da ateno e perda do controle emocional. b) rea temporoparietal: compreende todo o lbulo parietal inferior (giros supramarginal e angular e margens do sulco temporal superior e parte do lbulo parietal superior). Situa-se entre as reas secundrias auditiva, visual e somestsica, funcionando como centro que integra informaes recebidas dessas trs reas. Funcionalmente importante para a percepo espacial. c) reas lmbicas: compreendem o giro do cngulo, o giro para-hipocampal e o hipocampo. Essas reas, relacionadas principalmente com a memria e o comportamento emocional, juntamente com outras estruturas subcorticais integram o sistema lmbico.
- 15 -
2.1.2. Ncleos da Base
So tambm conhecidos como gnglios da base, porm essa nomenclatura errada, uma vez que fundamentalmente, gnglios so estruturas encontradas fora de cavidades sseas. Os ncleos da base constituem-se de: estriado, globo plido (segmentos internos e externos), ncleo subtalmico e substncia negra (parte compacta e reticulada). O estriado pode ser dividido em ncleo caudado, putamen e ncleo acumbens. Os dois primeiros ncleos so chamados de estriado dorsal e so separados pela cpsula interna. O ncleo acumbens tambm recebe o nome de estriado ventral. Funcionalmente, podemos dizer que as estruturas do corpo estriado participam da regulao do comportamento emocional por: 1. Se conectar com reas corticais do sistema lmbico, 2. processar informaes originadas em diversas reas do crtex cerebral, influenciando a atividade motora somtica (funo importante no planejamento motor), 3. influenciar reas nomotoras do crtex (rea pr-frontal, por exemplo) ligada exclusivamente a funes psquicas. A parte compacta da substncia negra formada em sua maioria por neurnios que utilizam a dopamina como neurotransmissor e possui importante conexo com o estriado. A degenerao de neurnios dopaminrgicos nesta rea causa diminuio de dopamina no estriado, provocando graves perturbaes motoras que caracterizam a sndrome de Parkinson, por exemplo.
Figura 6: Figura representando a localizao dos ncleos da base. (NETTER, Frank H.. Atlas de Anatomia Humana. 3ed. Porto Alegre: Artmed, 2004.)
- 16 -
2.1.3. Amgdala
O complexo amigdalar formado por mais de 10 ncleos e tem como papel central permitir que um organismo aprecie estmulos ambientais de significncia espcie-especfico, sendo ativada em situaes com significado emocional. Focos epilpticos nessa estrutura causam aumento da agressividade social, e a estimulao eltrica causa comportamento de fuga e defesa associadas agressividade. A memria, a ateno, o medo, a ansiedade, as reaes de defesa, os sistemas de recompensa, de reforo e de motivao, os comportamentos agressivos, maternal, sexual, ingestivo e do controle neuroendcrino esto de alguma forma direta ou indiretamente relacionados ao funcionamento da amgdala.
Figura 7: Imagem representativa da localizao da amgdala.
2.1.4. Formao Hipocampal Na superfcie medial do crebro, a formao hipocampal forma um anel que est situado dentro do crtex associativo e cerca o diencfalo e o mesencfalo. Trata-se de uma eminncia alongada e curva que no homem situa-se no assoalho do corno inferior dos ventrculos laterais. A formao hipocampal consiste de trs componentes: o subiculum, o hipocampo propriamente dito (ou corno de Ammon CA) e o giro denteado.
- 17 -
Figura 8:. Localizao do hipocampo. A formao hipocampal recebe sua maior aferncia do crtex entorrinal, regio localizada no giro parahipocampal adjacente formao hipocampal, que coleta informaes de outras regies associativas. Os neurnios eferentes da formao hipocampal so neurnios localizados no hipocampo e subiculum. Os axnios desses neurnios se agrupam em um feixe de fibras conhecido como frnix, esse se projeta para estruturas telenceflicas subcorticais e dienceflicas. O principal circuito que envolve a formao hipocampal conhecido como circuito tri-sinptico. Um esquema desse circuito est representado abaixo (figura 9).
Figura 9: Esquema representativo do circuito tri-sinptico. A formao hipocampal j foi relacionada regulao do comportamento emocional, uma vez que leses bilaterais do hipocampo em macacos resultam em aumento da agressividade desses animais. Posteriormente, foi visto que isto se dava graas leso tambm da amgdala, que relacionada ao comportamento emocional. Entretanto, a principal funo dessa regio a sua participao nos processos de memria, como visto no clssico caso do paciente H.M..
- 18 -
H.M. sofria de epilepsia intratvel (na ocasio); o foco epilptico situava-se no lobo temporal medial, bilateralmente. Ento, na tentativa de diminuir a intensidade e a frequncia das crises, removeu-se essa estrutura cirurgicamente. Isso resultou na remoo dos 2/3 anteriores dos hipocampos e das amgdalas, alm de outras pores corticais. Como esperado, a cirurgia apresentou xito quanto reduo da epilepsia; porm, H.M. deixou de ser capaz de manter informaes novas por mais de alguns minutos, nem reconhecia os mdicos que o acompanhava. Depois da remoo desta estrutura, H.M. era capaz de ler o mesmo jornal vrias vezes e surpreender-se sempre com as mesmas notcias. Entretanto, ele era capaz de aprender tarefas motoras bsicas, mas deveria ser lembrado de que ele sabia execut-las.
2.2. Diencfalo
O diencfalo, juntamente com o telencfalo, forma o crebro, e ocupa cerca de 80% da cavidade craniana. encoberto quase completamente pelos hemisfrios cerebrais, podendo ser visto apenas na face inferior do crebro. Compreende o tlamo, hipotlamo, epitlamo e subtlamo.
2.2.1. Tlamo
Na descrio de Angelo Machado (2006), os tlamos so duas massas volumosas de substncia cinzenta, de forma ovide, dispostas de cada lado, na poro ltero-dorsal do diencfalo. Funciona como um rel para as informaes sensoriais que se dirigem para o crtex cerebral, provenientes das regies mais caudais do SN. Os ncleos talmicos estabelecem conexo com o crtex cerebral por meio da cpsula interna. Funcionalmente os ncleos do tlamo (figura 10): 1) transportam informaes do tlamo para o giro do cngulo, participando da regulao da emoo; 2) transportam informaes motoras do globo plido e cerebelo para o lobo frontal; 3) recebem informaes sensoriais (dor, temperatura, presso e tato) dos lemniscos medial e espinal e envia projees para o giro ps-central; 4) projetam-se para o crtex de associao; 5) recebem fibras do lemnisco medial (rel de vias sensitivas) da sensibilidade da cabea;
- 19 -
6) alguns ncleos so inervados pelo crtex pr-frontal e recebem aferncias de outros ncleos talmicos e da amgdala; 7) alguns ncleos se projetam difusamente por todo o crtex e esto conectados ao SARA (sistema ativador reticular ascendente) - importante na regulao do sono e da viglia; 8) os ncleos geniculado lateral e medial medeiam informaes visuais e auditivas. As informaes auditivas provm dos colculos.
Figura 10: Localizao do tlamo, ncleos talmicos e suas conexes.
Afeces do tlamo, que podem ocorrer devido a leses em alguns vasos, resultam na chamada sndrome talmica. Esta sndrome acarreta em alteraes da sensibilidade. Uma delas o aparecimento de crises, denominadas dor central, que se caracteriza por uma dor pouco localizada e espontnea, que pode se irradiar a toda metade do corpo que se situa do lado oposto ao tlamo comprometido.
- 20 -
2.2.2. Subtlamo
Baseado nas descries de Angelo Machado (2006), o subtlamo uma estrutura de pequena extenso situada na parte posterior do diencfalo na transio com o mesencfalo limitando-se superiormente com o tlamo, lateralmente com a cpsula interna e medialmente com o hipotlamo. Devido sua localizao, algumas estruturas mesenceflicas estendem-se at o subtlamo, como o ncleo rubro, a substncia negra e a formao reticular, constituindo esta a chamada zona incerta do subtlamo. Entretanto, apresenta algumas formaes cinzentas e brancas que lhe so prprias, sendo a mais importante o ncleo subtalmico que tem conexo com o globo plido pelo circuito plido-subtlamo-palidal, importante para a regulao da motricidade somtica. Leses nesta estrutura provocam o hemibalismo, sndrome caracterizada por movimentos anormais das extremidades.
2.2.3. Epitlamo
Localiza-se na parte superior e posterior do diencfalo e contm formaes endcrinas e no-endcrinas. A formao endcrina mais importante a glndula pineal; as formaes noendcrinas so os ncleos das habnulas, a comissura das habnulas, as estrias medulares e a comissura posterior. A glndula pineal importante, pois est relacionada produo e liberao de melatonina, um hormnio envolvido com os ritmos circadianos (especialmente o sono e a viglia).
2.2.4. Hipotlamo
Estrutura constituda fundamentalmente por substncia cinzenta agrupada em ncleos. O frnix, estrutura pertencente ao circuito de Papez envolvido nos mecanismos da memria e das emoes, divide o hipotlamo em uma rea medial e outra lateral. Possui conexes amplas que incluem o sistema lmbico (regulao do comportamento emocional), a rea pr-frontal (comportamento emocional), a hipfise, as vias sensoriais (provenientes das vsceras), neurnios monoaminrgicos (noradrenrgicos da formao reticular e serotoninrgicos, dos ncleos da rafe). Como quase todas as suas funes se relacionam com a homeostase, tem um papel regulador sobre o SNA e o sistema endcrino, alm de controlar vrios processos motivacionais importantes para a sobrevivncia do indivduo e da espcie (fome, sede, sexo, etc). Suas principais funes so: regulao da temperatura corporal, regulao do comportamento
- 21 -
emocional, regulao do sono e da viglia, regulao da ingesto de alimentos e gua, regulao da diurese, regulao do sistema endcrino, gerao e regulao de ritmos circadianos.
Figura 11: Figura ilustrando a localizao e a diviso do hipotlamo
2.2.5. Hipfise
Conhecida anteriormente como glndula pituitria, a hipfise uma estrutura localizada em uma cavidade ssea, a sela trcica. Produz hormnios de ampla atuao no organismo e comandada pelo hipotlamo. Fisiologicamente, pode ser dividida em neuro e adenoipfise. A primeira de origem nervosa, e a segunda, epitelial. A neuroipfise ligada ao hipotlamo pelo infundbulo e secreta hormnios produzidos por este. Os corpos celulares dos neurnios se encontram no hipotlamo, e os axnios dirigem os hormnios neuroipfise. responsvel pela secreo de hormnio antidiurtico (ADH) e ocitocina. A adenoipfise constituda de epitlio glandular e tambm recebe ordens do hipotlamo, atravs do sistema de vasos porta hipotalmico-hipofisrios. Esta regio da hipfise responsvel pela secreo de prolactina, hormnio do crescimento (GH), hormnio folculo estimulante (FSH), hormnio luteinizante (LH), hormnio estimulante da tireide, hormnio adrenocorticotrfico e endorfinas. A estreita relao entre hipfise e hipotlamo evidencia-se ao observarmos que, em geral, distrbios endcrinos cursam com sintomas psiquitricos e vice-versa.
- 22 -
Figura 12: Figura representando a localizao da hipfise (pituitria) no encfalo.
2.3. Tronco enceflico
O tronco cerebral, anatomicamente localizado entre o diencfalo e a medula espinhal e ventralmente ao cerebelo, compreende estruturas originadas de trs vesculas embrionrias: mesencfalo, metencfalo e mielencfalo. O mesencfalo no se subdivide como os demais, e continua a ser denominado mesencfalo; o metencfalo, por sua vez, origina a ponte e o cerebelo; o mielencfalo origina o bulbo. O tronco enceflico composto por feixes que atuam como passagem de informaes sensitivas e motoras. Estes feixes podem originar-se do prprio tronco, enquanto outros somente o atravessam. Dos 12 pares de nervos cranianos, 10 passam pelo tronco e tem maior complexidade que a medula, pois possui centros que interagem com outras estruturas do sistema nervoso central com diversas funes.
Figura 13: Localizao e diviso do tronco enceflico. (NETTER, Frank H.. Atlas de Anatomia Humana. 3ed. Porto Alegre: Artmed, 2004).
- 23 -
Sua estrutura interna semelhante a da medula, onde a substncia cinzenta fica localizada internamente e a substncia branca externamente. Contudo, a substncia cinzenta do tronco fragmentada transversal e longitudinalmente dando origem a ncleos. Alguns destes ncleos, quando em conjunto formam a substncia cinzenta homloga (apresenta a mesma relao com os nervos cranianos que a substncia cinzenta da medula com os nervos espinhais) ou substncia cinzenta prpria (sem relao com os nervos cranianos). Existem ainda, em toda extenso do tronco, um emaranhado de clulas e fibras nervosas com uma estrutura intermediria entre a substncia cinzenta e branca, chamada de Formao reticular. A substncia branca caracterizada pela passagem dos tratos ascendentes, descendentes e de associao, que trazem ou levam informaes do crtex para medula espinhal. O tronco enceflico se interpe entre a medula e o diencfalo, situando-se ventralmente ao cerebelo e dividido em 3 regies: bulbo, ponte e mesencfalo.
2.3.1. Mesencfalo
Estende-se da ponte at o diencfalo e o terceiro ventrculo, possuindo vrios ncleos de nervos cranianos. Suas principais estruturas so: 1) Substncia Cinzenta Periaquedutal cuja parte dorsal integra comportamentos defensivos envolvidos no medo e ansiedade e sua parte ventral, onde atuam mecanismos de controle da dor, 2) Colculos superiores, relacionados com rgos da viso e inferiores, com rgos da audio, 3) Ncleo Rubro que participa do controle da motricidade somtica e 4) Substncia Negra, cuja maioria dos neurnios dopaminrgico; possui importante conexo com o estriado. A degenerao de neurnios nesta rea causa diminuio de dopamina no estriado, provocando graves perturbaes motoras que caracterizam a sndrome de Parkinson. Na regio anterior do ramo longitudinal da cruz aloja-se o corpo pineal (formao endcrina responsvel pela secreo de melatonina, com ao antigonadotrfica e que participa dos ritmos circadianos), que, entretanto, pertence ao diencfalo. Cada colculo se liga a uma pequena eminncia oval do diencfalo, o corpo geniculado, atravs de um feixe superficial de fibras nervosas que constitui o seu brao.
2.3.2. Cerebelo
O cerebelo faz parte do sistema supra-segmentar e deriva da parte dorsal do metencfalo. situado dorsalmente ao bulbo e ponte, contribuindo para a formao do tecto do quarto ventrculo.
- 24 -
Seu crtex envolve o corpo medular do cerebelo. Fisiologicamente se difere do crebro por funcionar sempre em nvel involuntrio e inconsciente. Seus aspectos funcionais englobam a manuteno do equilbrio e postura, controle do tnus muscular, controle dos movimentos voluntrios (planejamento do movimento e correo dos movimentos) e aprendizagem motora (ao executarmos vrias vezes a mesma atividade motora, esta passa a ser cada vez mais rpida e eficiente). Os principais sintomas de leses no cerebelo so a incoordenao dos movimentos (ataxia) manifestando-se principalmente nos membros (marcha atxica) ou na articulao das palavras (voz arrastada), a perda do equilbrio e a diminuio da musculatura esqueltica (hipotonia).
Figura 14: Ilustrao do Cerebelo (NETTER, Frank H.. Atlas de Anatomia Humana. 3ed. Porto Alegre: Artmed, 2004.)
2.3.3. Ponte A ponte fica situada entre o bulbo e o mesencfalo e sua caracterstica mais marcante uma grande massa ovide na superfcie ventral do tronco enceflico: a ponte basal. Assim como no bulbo, na ponte tambm existem ncleos de nervos cranianos sendo que leses nesses ncleos envolvem alterao da sensibilidade da face, alterao da motricidade da musculatura mastigadora ou mmica, tontura, e alteraes do equilbrio associadas paralisia ou perda da sensibilidade motora do tronco e membros. Os ncleos pontinos da ponte basal transmitem informao oriunda do crtex cerebral, para o cerebelo.
- 25 -
A ponte funciona como estao para as informaes provenientes dos hemisfrios cerebrais e que se dirigem para o cerebelo. Na transio entre o bulbo e a ponte est localizado o locus coeruleus, principal fonte de inervao noradrenrgica no SNC, que possui importante papel no controle emocional e no ciclo sono-viglia.
2.3.4. Bulbo
O bulbo, bulbo raqudeo ou ainda medula oblonga, a parte menor e mais caudal do tronco enceflico, continua inferiormente com a medula espinal e, superiormente com a ponte. Conecta a regio menos diferenciada do SNC, a medula espinal, com as regies mais diferenciadas do encfalo. Apesar de ser uma parte relativamente pequena do SNC percorrido por um grande nmero de tratos motores e sensitivos, possuindo, ainda, ncleos importantes, principalmente ncleos dos nervos cranianos. Leses nessa rea causam dificuldades de deglutio, alteraes na fonao e nos movimentos da lngua. A leso de determinadas fibras pode causar perda da sensibilidade do tronco e membros. Sua formao reticular abriga os centros do vmito, respiratrio e vasomotor, esses ltimos importantes para o controle do ritmo respiratrio, cardaco e presso arterial. Na regio posterior do bulbo encontramos os fascculos grcil e cuneiforme, constitudos por fibras nervosas ascendentes, provenientes da medula, que terminam em duas massas de substncia cinzenta, os ncleos grcil e cuneiforme, envolvidos na transmisso sensitiva dos impulsos da propriocepo consciente, tato epicrtico e sensibilidade vibratria que se dirigem para o tlamo. Encontramos ainda, as chamadas pirmides bulbares que so atravessadas pelo trato crticoespinal (ou trato piramidal) ligando as reas motoras do crebro aos neurnios motores da medula.
2.3.5. Formao Reticular
uma agregao mais ou menos difusa de neurnios de tamanhos e tipos diferentes, separados por uma rede de fibras nervosas que ocupa a parte central do tronco enceflico, preenchendo todo o espao que no preenchido pelos tractos, fascculos e ncleos de estrutura mais compacta. Tem importante papel na ativao do crtex cerebral. Embora no tenha estrutura homognea.
- 26 -
A formao reticular possui conexes amplas e variadas que, alm de receber impulsos que entram pelos nervos cranianos, mantm relaes nos dois sentidos com o crebro, cerebelo e medula espinal. Funcionalmente, influencia quase todas as funes controladas pelo SNC: atividade eltrica cortical (sono e viglia), motricidade somtica, sistema nervoso autnomo, controle eferente da sensibilidade, controle neuroendcrino e integrao de reflexos (centro respiratrio e vasomotor).
2.4. Medula Espinhal
A medula espinal uma massa cilindride, ligeiramente achatada no sentido nteroposterior, de tecido nervoso, situada dentro do canal vertebral sem, porm, ocup-lo completamente, medindo aproximadamente 45 cm em um humano adulto. Limita-se cranialmente com o bulbo (ao nvel do forame magno do osso occipital). Seu limite caudal tem importncia clnica situando-se, no humano adulto, geralmente na L2. A medula termina afilandose para formar um cone (cone medular) que continua com um delgado filamento menngeo, o filamento terminal. A medula espinhal apresenta duas dilataes, as intumescncias cervical e lombar, situadas nas regies cervical e lombar, respectivamente, correspondendo s reas em que grossas razes nervosas fazem conexes com a medula, formando os plexos braquial e lombossacral, destinados inervao dos membros superiores e inferiores. Na medula, a substncia cinzenta localiza-se por dentro da branca, que composta basicamente por fibras mielnicas que sobem e descem na medula, e apresenta forma de um H. Da medula espinal emergem os 31 pares de nervos espinais.
- 27 -
Figura 15: Estrutura da medula espinhal. (NETTER, Frank H.. Atlas de Anatomia Humana. 3ed. Porto Alegre: Artmed, 2004.)
A medula no apenas um meio em que as informaes sensoriais e motoras chegam ao crebro, mas tambm de informaes do crebro para os rgos, alm de poder desencadear processos de forma autnoma como os processos de reflexos simples. Levando-se em considerao as funes motoras, a medula espinhal pode ser organizada da seguinte maneira: por onde entra e por onde sai a informao. Na raiz posterior onde entra j a sada ocorre pela raiz anterior da medula e tem como principais alvos os msculos esquelticos. Sendo assim, como todos os nervos que emergem da medula espinhal se unem com os que saem, formam um Y, em que sua raiz dorsal entra no corno posterior, com informao sensitiva e em sua raiz ventral, entra no corno anterior, com informao motora. Os neurnios motores da raiz anterior se diferenciam em alpha e gamma. Os alfas so responsveis pela excitao de unidades motoras, j os gamas, pela inervao de fibras. Interneurnios so tambm neurnios bastante presentes no H medular, e so responsveis por funes integrativas. Fazem o processamento de informaes do momento em que estas chegam at os momentos em que saem da medula.
- 28 -
Arco Reflexo Simples Um desequilbrio transitrio do ambiente pode estimular neurnios sensoriais e produzir um reflexo motor (como o reflexo patelar), atravs de um neurnio motor. Esse tipo de reflexo constitui um grupo especial de reflexos denominados de espinhais, ou seja, todo o comportamento restrito a conexes com a medula e no chega ao encfalo. So geralmente mediados por circuitos monossinpticos, nos quais os neurnios sensoriais esto diretamente conectados aos neurnios motores. O reflexo patelar e seus componentes anatmicos esto representados na figura 16. Uma leve batida do tendo patelar, que liga o msculo quadrceps femoral patela, produz um reflexo rpido de estiramento (como um chute), atravs da contrao do msculo da coxa, quadrceps femoral, e relaxamento dos msculos flexores opostos da parte de trs da coxa.
Figura 16: O reflexo patelar um exemplo de um sistema reflexo monossinptico, um
comportamento simples controlado por conexes diretas entre os neurnios sensoriais e motores. Bater levemente na patela com um martelo de reflexo causa um puxo no tendo do quadrceps femoral. Quando o msculo estirado, reagindo ao puxo no tendo, a informao a respeito dessa mudana no msculo transmitida por neurnios aferentes (sensoriais) para o Sistema Nervoso Central. Na medula espinal, os neurnios sensoriais agem diretamente nos neurnios motores extensores que contraem o quadrceps. Alm disso, os neurnios sensoriais agem indiretamente, por meio de interneurnios, para inibir os neurnios motores flexores, que de outra forma contrairiam os msculos opostos (squios tibiais). Essas aes combinam-se para produzir o comportamento reflexo. Nesse desenho esquemtico cada neurnio motor extensor e flexor representa uma populao de muitas clulas (divergncia neuronal).
Os corpos celulares dos neurnios sensoriais mecanoceptores envolvidos no reflexo patelar esto agrupados prximos medula espinal em um gnglio da raiz dorsal. Um ramo do axnio do neurnio sensorial vai at o msculo quadrceps femoral e o outro se localiza centralmente da medula espinhal. O ramo da medula espinhal faz sinapse com os neurnios motores que tem seus corpos celulares localizados no gnglio da raiz ventral. O neurnio motor inerva o msculo quadrceps femoral e controla a contrao muscular. Esse neurnio tambm faz conexes com
- 29 -
neurnios controladores dos msculos antagnicos contrao em questo, facilitando o movimento reflexo de estiramento (figura 16).
- 30 -
REFERNCIAS BIBLIOGRFICAS
Bear, Mark F. Neuroscience: exploring the brain. Mark F Bear, Barry W Connors, Michel A Paradiso. 3a edio. Lippincott Williams & Wilkins, 2007 Kandel, Eric R. principles of neural science. Eric R Kandel, James H Schwartz, Thomas M Jessel. 4a edio. McGraw-Hill Companies, 2000 Machado, Angelo BM. Neuroanatomia Funcional. 2a edio. Editora Atheneu, 2005 Martin, John H. Neuroanatomy: text and atlas. 3a edio. McGraw-Hill Companies, 2003 Netter, Frank H.. Atlas de Anatomia Humana. 3ed. Porto Alegre: Artmed, 2004.
- 31 -
NEUROFISIOLOGIA
1. A MEMBRANA CELULAR E SEUS TRANSPORTES
Para o entendimento do funcionamento das sinapses e das propriedades eltricas dos neurnios, necessrio se levar em conta o funcionamento da membrana celular. Esta formada por uma bicamada lipdica que no se difunde com os meios intra e extracelular. Essa uma caracterstica importante para a separao desses meios e manuteno da constncia do meio interno, fundamental para o funcionamento celular. Apesar da estrutura lipdica da membrana, protenas tm capacidade de atravess-la e possibilitar a passagem de molculas de um meio para outro. Estas so chamadas de protenas de transporte e so divididas conforme a forma que executam essa funo. As que possuem canais aquosos no seu interior e possibilitam a passagem de determinados ons e molculas so chamadas protenas de canal, j as que tm a caracterstica de se acoplarem molcula a ser transportada e mudar sua conformao para a passagem, so as protenas carreadoras. Mesmo se diferindo em alguns aspectos, essas protenas so seletivas ao que transportam, respondendo a necessidades celulares.
Figura 1: Mecanismos bsicos de transporte celular. (Guyton, 9 edio)
O transporte pela membrana seja atravs da camada bilipdica, ou pelas protenas, pode ocorrer basicamente por: transporte ativo ou passivo (difuso) (figura 1). - Difuso: Transporte sem gasto de energia, ou seja, passivo. Pode ser subdividido em 2, difuso simples ou facilitada. o Difuso simples: movimento molecular e inico atravs da membrana sem a necessidade de protenas. A velocidade depende das concentraes da substncia nos meios intra e extracelular, nmero de locais na membrana possveis para a passagem e a velocidade que a substncia consegue passar pela membrana. Este ltimo, dependente da carga e
- 32 -
do tamanho da substncia, principalmente. Esse tipo de transporte pode ocorrer atravs do prprio interstcio da membrana ou pelos canais inicos. a forma de passagem de substncias lipossolveis, j que apenas estas substncias tm caractersticas semelhantes membrana. o Difuso facilitada: H a necessidade de interao da molcula ou on com protenas de membrana, que muda sua conformao para permitir o transporte (figura 2).
Figura 2: Funcionamento da protena carreadora em difuso facilitada. (Guyton, 9 edio)
Pela necessidade de acoplamento entre as molculas e alterao conformacional da protena transportadora, este processo mais demorado do que a difuso simples. Alm disso, como h a necessidade de protenas carreadoras especficas, no s a quantidade da molcula a ser transportada influencia, mas tambm a quantidade dessas protenas. A diferena entre difuso simples e facilitada em termos prticos pode ser representada pela figura 3.
Figura 3: Velocidade de difuso pela concentrao da molcula a ser transportada (Guyton, 9 edio)
- 33 -
Entre as molculas mais importantes transportadas pela difuso facilitada, esto: glicose e aminocidos. A gua, substncia mais abundante difundida atravs da membrana, sofre interferncia de sua prpria concentrao para sua movimentao. Esse fenmeno chamado osmose e ocorre nas duas direes (do meio interno para o externo e vice-versa). Geralmente, h um balano fino entre a entrada e sada de gua, o que faz com que a clula permanea do mesmo tamanho, mas em algumas condies, pode haver murchamento ou intumescimento celular.
Figura 4: Representao do princpio da osmose. (Guyton, 9 edio)
A figura acima representa o comportamento da gua. Na situao inicial, uma membrana impermevel ao soluto separa duas solues de concentraes diferentes. Na soluo hipertnica, a concentrao de gua em relao ao soluto menor, o que faz com que por osmose, gua da soluo hipotnica atravesse a membrana para que as concentraes se assemelhem. Mesmo depois que as concentraes ficam iguais, a difuso de gua continua, porm nas duas direes. Esse tipo de comportamento chamado equilbrio dinmico. A osmose pode ser lentificada ou at cessada se houver alguma resistncia passagem de gua. O aumento de presso do lado hipertnico capaz de lentificar ou cessar a passagem da gua. Esta presso chamada presso osmtica, ou seja, presso que cessa a osmose. As foras que permeiam o transporte passivo so denominadas de foras eletroqumicas, ou seja, dependentes das concentraes, potencial eltrico e presso osmtica. Entretanto, pode ocorrer grande necessidade de alguma substncia no meio intracelular, mesmo que o extracelular no o oferea em grandes quantidades. Nestes casos, h a necessidade de gasto energtico para um transporte independente das foras eletroqumicas. - Transporte ativo: dependente da utilizao de energia e importante para os ons potssio, sdio, clcio, hidrognio, cloreto, aminocidos, entre outros. Para todos os casos, h a utilizao de protenas carreadoras. dividido em primrio e secundrio, conforme fonte de energia utilizada. No primeiro caso, a energia vem diretamente da desfosforilao do ATP ou qualquer outro composto fosfatado rico em energia.
- 34 -
Um exemplo a bomba de sdio e potssio, que faz o transporte de ons sdio para fora e potssio para dentro da clula. Esta ao contribui para o estabelecimento de um potencial eltrico negativo no interior celular.
Figura 5: Esquema do funcionamento da bomba de sdio e potssio. H a quebra do ATP em ADP e liberao de fsforo inorgnico na passagem de 3 ons Na + para o meio extracelular enquanto h a internalizao de 2 ons K+ (Guyton, 9 edio)
No caso do transporte ativo secundrio (ou cotransporte), a energia provm de gradientes inicos formados a partir do transporte ativo primrio. Um exemplo quando h o transporte ativo primrio de Na+ para fora da clula, o que provoca um aumento da concentrao deste on no meio extracelular. A diferena de concentraes entre os meios intra e extracelulares faz com que haja uma fora do Na+ de entrada na clula. Esta fora tal que consegue levar outros ons juntamente, de carona. H casos tambm em que o cotransporte ocorre em direes opostas.
2. TRANSPORTE E CARGAS NA MEMBRANA PLASMTICA
A diferena nas concentraes de ons e molculas entre as faces da membrana o principal determinante do potencial de repouso, que uma desigualdade eltrica entre os meios intra e extracelulares. O transporte desses ons e molculas de um lado para o outro da membrana o que causa variaes nesse potencial eltrico e que tem grande importncia na transferncia de informaes entre neurnios (sinapses) e dentro deles. Considerando uma clula hipottica em que no haja transporte ativo e a difuso seja a responsvel pela homeostase eletroqumica. Na figura 6, h grande concentrao de ons K+
- 35 -
dentro da clula em relao ao meio externo e no h diferena eltrica entre os meios (situao A). Alm disso, a membrana permevel apenas a este on.
Figura 6: Representao da distribuio dos ons potssio. Por causa da diferena entre as concentraes, o K+ tende a se dirigir ao meio externo. Como o K+ um ction, sua sada faz com que o meio externo comece a apresentar uma positividade eltrica e o interno, como manteve os nions, uma negatividade (situao B). A passagem do K+ para o meio externo contnua at que a quantidade de cargas positivas deste lado comece a exercer uma fora eltrica contrria sada do K +. Isso ocorre por ao da lei em que os opostos se atraem e semelhantes se repelem. Essa fora eltrica exercida contra a passagem do K+ chega a bloquear seu efluxo, apesar de sua concentrao continuar maior no meio interno. Ou seja, a fora eltrica se iguala qumica e a difuso cessa. basicamente dessa forma que os ons e molculas proporcionam um potencial de membrana, porm, biologicamente, so muitas as substncias que influenciam esse processo.
3. TIPOS DE SINAPSES
Sinapse, ou comunicao intercelular, a responsvel pela passagem de informaes de uma parte do SN para outra e essa capacidade o que distingue o neurnio das demais clulas. O termo sinapse provm da palavra grega conectar e foi introduzido pelo fisiologista britnico Charles Sherrington no final do sculo XIX. H pouco tempo, considerava-se que uma sinapse fosse formada basicamente por dois elementos: o terminal pr-sinptico e a ps-sinapse, ambos neuronais. Em algumas sinapses, o que ocorre basicamente isto, estando ambos separados apenas por um espao virtual denominado fenda sinptica. As sinapses podem ser classificadas quanto sua natureza, em eltricas ou qumicas, e quanto sua funo em excitatrias ou inibitrias.
- 36 -
Sinapses eltricas: so morfologicamente simtricas (sem particularidades morfolgicas que permitam a distino entre os elementos pr e ps-sinpticos, vistos por microscopia eletrnica), e a transferncia de informaes bidirecional. H reas de justaposio entre as membranas pr e ps-sinpticas em que se encontram as junes gap. Estas junes se apresentam como poros que fazem a conexo clula-clula e permitem a passagem livre de ons e pequenas molculas do citoplasma de uma clula para outra. A importncia da sinapse eltrica se d principalmente pela transferncia de informaes de maneira bidirecional e transmisso rpida de sinais, onde no existe perda ou atraso na passagem da informao. Dessa forma, responsvel pela sincronizao da atividade eltrica em uma grande populao neuronal. Sinapses qumicas: possuem assimetria morfolgica, com caractersticas distintas entre os elementos pr e ps-sinpticos. O terminal pr-sinptico a poro final do axnio levemente intumescido e com maior nmero de mitocndrias, alm de uma variedade vesculas. Nesse tipo de sinapse no h contato entre as membranas das clulas pr e ps-sinpticas, alm de haver um maior espao entre elas (fenda sinptica). De maneira geral, estas vesculas esto agrupadas nos arredores do terminal axnico (figura 7), e no ponto de maior proximidade com a membrana da clula ps-sinptica. No elemento ps-sinptico no existe a presena de vesculas prximas membrana, mas, frequentemente, h uma estrutura altamente eletrondensa, denominada densidade pssinptica. Acredita-se que a funo desta estrutura seja auxiliar a ancoragem de receptores de neurotransmissores na membrana ps-sinptica.
Figura 7: Micrografia eletrnica mostrando a fuso entre vesculas sinpticas e a membrana plasmtica de um neurnio pr-sinptico. A seta indica a abertura inicial do poro da vescula para a liberao do neurotransmissor. A ponta de seta mostra uma vescula fundida a membrana. A chave ilustra uma regio da bicamada fosfolipdica do neurnio. Barra de escala 5 m.
Neste tipo de sinapse a transferncia de sinais apenas unidirecional, sempre no sentido do neurnio pr para o ps-sinptico. A sinapse qumica sofre um atraso, por possuir um espao entre as membranas e ter a transmisso da informao dependente de neurotransmissores e receptores.
- 37 -
4. SINALIZAO NEURONAL
4.1. Potencial de Membrana
O fluxo de informao inter e intraneurnios realizado por meio de sinais qumicos e eltricos. Os sinais eltricos so todos produzidos por variaes no fluxo ons atravs da membrana, o que gera uma diferena de potencial eltrico entre as faces desta. O controle do fluxo dado por diferentes canais inicos inseridos na membrana plasmtica das clulas. Existem dois grandes tipos de canais inicos: - Canais que permanecem constantemente abertos e no so influenciados por fatores externos. So importantes para a manuteno do potencial de repouso da membrana; - Canais que se encontram fechados na condio de repouso, cuja abertura e fechamento dependem de fatores externos, tais como: presena de um ligante, alteraes no potencial de membrana, ou seu estiramento. No repouso, o neurnio tem excesso de cargas positivas na face externa da membrana e excesso de cargas negativas na face interna. Essa separao de cargas origina uma diferena de potencial eltrico: potencial de membrana (Vm) que o potencial intracelular (V int) subtrado do potencial extracelular (Vext). No repouso, este potencial de membrana tem valores de 60 a 70 mV, e denominado potencial de repouso da membrana. As sinalizaes eltricas que ocorrem nos neurnios resultam de breves alteraes do potencial de repouso da membrana, e essas alteraes podem ser despolarizantes (diminuindo a separao de cargas atravs da membrana) ou hiperpolarizantes (aumentando a separao de cargas atravs da membrana) dependendo do fluxo de ctions e/ou nions para dentro ou para fora da clula. O potencial de repouso da membrana gerado e mantido pela diferena de concentrao inica entre os meios intra e extracelular. Dos quatro tipos mais abundantes de ons encontrados dentro e fora da clula, o Na+ e o Cl- so mais concentrados no meio extracelular; j o K+ e os nions orgnicos (A-), como aminocidos e protenas, esto mais concentrados no meio intracelular (tabela 1). Esta diferena de concentrao inica gera dois tipos de foras (1) a fora qumica, de difuso, dada pela diferena de concentrao de ons e (2) a fora eltrica, de presso eletrosttica, dada pela diferena de cargas positivas e negativas dispostas nos dois lados da membrana. A resultante destas duas foras - a fora eletroqumica - tem componentes qumicos e eltricos provenientes da fora de difuso e da fora eletrosttica, respectivamente.
- 38 -
Tabela 1: ons e potenciais de membrana. No caso do on K+, por exemplo, a fora de difuso atua no sentido de efluxo do on, pois o mesmo encontra-se mais concentrado no meio intracelular do que no extracelular. Contudo, a fora eltrica atua no sentido oposto, mantendo o potssio, um ction, no interior da clula que predominantemente negativo. A resultante entre estas foras o potencial de equilbrio do K +. No caso do Na+, a situao diferente. Sua concentrao maior no meio externo do que no meio interno, sendo assim, tende a fluir para dentro da clula. Alm disso, o Na + tambm direcionado para o interior da clula pelo gradiente eltrico (j que o meio intracelular mais negativo), ento deveria ocorrer o influxo de Na+, porm isso no acontece de forma expressiva, devido permeabilidade seletiva da membrana. O fluxo de ons atravs da membrana produto da fora eletroqumica em funo da permeabilidade da membrana ao on. Na clula em repouso (Vm = VR); h poucos canais de Na+ abertos, resultando em um fluxo de Na+ pequeno, ainda que as foras qumica e eltrica o forcem para o interior da clula. Por apresentar grande permeabilidade (cerca de 100 vezes maior do que o do Na +), ocorre efluxo do K+, apesar da fora eltrica ser exercida para mant-lo no interior da clula. Resumindo, como as concentraes so bastante diferentes entre os meios intra e extracelulares, o fluxo desses ons tenderia ao equilbrio, portanto necessrio que haja uma fora atuante no sentido contrrio ao gradiente, com a finalidade de manter essas diferenas de concentrao intra e extracelular. Esta funo exercida por transportadores ativos. No caso dos ons Na+ e K+, o transportador mais importante para a manuteno da homeostase, so as bombas de Na+/K+ na membrana. A bomba Na+/K+ atua transportando, contra o gradiente, 3 molculas de Na + e 2 molculas de K+, utilizando uma molcula de ATP. Ou seja, 3 molculas de Na + so transportadas para o meio extracelular, enquanto 2 molculas de K+ para o meio intracelular. Sendo assim, seu papel fundamental na manuteno do potencial de repouso da membrana, j que os gradientes de concentrao para Na+ e K+ devem permanecer constantes. Embora os fluxos de Na+ e K+ definam o potencial de repouso da membrana, este influenciado pelas concentraes de mais ons e respectivas permeabilidades.
- 39 -
4.2. Potencial de Ao (PA)
O potencial de membrana, na condio de repouso, mantido por um equilbrio dinmico entre os componentes dos meios intra e extracelulares. O Na + e o K+ so os principais ons na manuteno deste potencial. No caso do potencial de ao, uma alterao no potencial de repouso leva transmisso de informaes por sinais eltricos ao longo do neurnio. O potencial de ao iniciado quando um neurnio recebe um estmulo que despolariza sua membrana. Esse estmulo deve atingir um limiar mnimo para que o PA seja gerado (cerca de 55mV). Este estmulo suficiente para alterar a diferena de potencial de membrana e estimular canais voltagem dependentes de Na+ e K+, causando a abertura destes. A necessidade desse limiar mnimo (limiar de disparo) para a deflagrao do PA d sua caracterstica tudo ou nada. Ou seja, um estmulo inferior no vai faze r com que haja alterao nos canais, ainda que seja prximo do limiar, e um estmulo superior no vai fazer com que haja a gerao de um potencial mais forte ou mais de um potencial. De maneira geral, os neurnios sofrem uma srie de estmulos, mas que no so suficientes para a deflagrao do potencial de ao. Estes pequenos estmulos fazem parte da dinmica normal do neurnio, e no tem consequncias na eletrofisiologia neuronal, ou seja, sendo o limiar de disparo mnimo de -30mV e o estmulo, de -25mV, o potencial no deflagrado. Os canais de Na+ voltagem dependentes respondem rapidamente despolarizao e se abrem, permitindo o influxo de Na+ at que o potencial de membrana seja invertido e alcance cerca de +30mV. Quando esse potencial alcanado, os canais de Na+ so fechados e o influxo cessa. Os canais de K+ tambm so ativados no incio da despolarizao da membrana, porm sua ativao lenta, o que faz com que sua abertura seja depois que os canais de Na+ j comearam a se fechar. O papel do K+ no potencial de ao principalmente o retorno ao repouso. Essa variao no potencial da membrana representada na figura 8.
- 40 -
Figura 8: Esquema representativo de um potencial de ao.
Nesse grfico, pode-se observar uma fase inicial de ascenso (1) que representa a despolarizao. Aps atingir o pico mximo de despolarizao, ocorre um declnio do potencial de membrana no sentido de reestabelecer o potencial de repouso (2). Depois disso, a clula ainda sofre uma pequena hiperpolarizao (3) - aumenta a negatividade do potencial de membrana. A fase de hiperpolarizao est relacionada ao efluxo de K+ aps o fechamento dos canais de Na+. Os canais de K+ agem de uma forma mais lenta, continuando a agir aps o potencial de ao j ter terminado. Essa fase importante na passagem unidirecional da informao, uma vez que a rea hiperpolarizada no responde a outro limiar at voltar ao potencial de repouso. Nesta fase, ocorre um perodo refratrio breve, dividido em duas fases: um perodo refratrio absoluto que ocorre imediatamente aps o potencial de ao e um perodo refratrio relativo que se segue ao anterior. Na primeira fase impossvel excitar uma clula, independentemente do quo intenso o estmulo despolarizante aplicado. O mesmo no ocorre no perodo seguinte porque se o estmulo aplicado for intenso, ento a clula capaz de deflagrar um novo potencial de ao. O perodo de refrao dura por milissegundos e consequncia dos canais de Na+ ainda inativados e os de K+ ainda abertos. Em geral, os mecanismos bsicos para gerao do potencial de ao so os mesmos em todos os neurnios, embora os diferentes tipos de canais inicos proporcionem distintos graus de excitabilidade da membrana.
- 41 -
4.3. Propagao do Potencial de Ao
Gerado o potencial de ao, necessrio que ele se propague para que cumpra a funo de transmisso de informao. Em geral, o potencial de ao produzido em um determinado ponto da membrana excita as regies adjacentes a este ponto. Como consequncia, os canais de Na+ nestas regies abrem-se imediatamente e o potencial de ao vai sendo propagado. Este processo acontece repetidamente, sendo que a despolarizao trafega ao longo da extenso de toda a fibra nervosa. A transmisso do processo de despolarizao ao longo da fibra nervosa ou muscular denominada de impulso nervoso ou muscular, respectivamente. Para que haja a propagao do potencial de ao gerado, os neurnios apresentam propriedades eltricas passivas. So elas: resistncia e capacitncia da membrana e resistncia axial intracelular ao longo dos dendritos e do axnio. Esta propagao de sinal entre neurnios influenciada principalmente por 2 constantes: - Constante de espao: Perda de sinal ao longo do comprimento axonal de maneira que quanto maior for resistncia da membrana (isto , quanto menos canais inicos de repouso tiver a membrana), maior ser esta constante, portanto, mais distante se propagar um dado sinal eltrico. - Constante de tempo: Determina o quo rapidamente o potencial varia ao longo do tempo, independe do tamanho celular, porm de sua composio. Esta relacionada capacidade de somao de estmulos. Supondo que dois estmulos despolarizantes atinjam uma nica clula, se esse neurnio tiver uma alta constante de tempo pode ocorrer a somao temporal, que resulta em uma maior durao do potencial excitatrio; se o mesmo neurnio tiver uma alta constante de espao, por sua vez, ocorre a somao espacial que permite que os potenciais sejam somados, conferindo uma maior amplitude do sinal. Dessa forma, a constante de tempo fornece a durao do potencial excitatrio, enquanto que a constante de espao est relacionada sua amplitude. O potencial de ao se propaga rapidamente pela regio revestida com mielina (devido baixa resistncia) e diminui de velocidade ao chegar a cada ndulo de Ranvier (devido alta resistncia). Sendo assim, o potencial de ao parece saltar rapidamente de nodo para nodo. Esse comportamento, em axnios mielinizados, possui o nome de conduo saltatria. A conduo saltatria tambm metabolicamente favorvel, pois menos energia dispendida na manuteno da bomba Na+/K+. Esse o mecanismo bsico da conduo dos sinais eltricos ao longo do neurnio. Geralmente, quando o potencial de ao chega regio dos terminais sinpticos, esses sinais eltricos so convertidos em sinais qumicos pela liberao dos neurotransmissores na fenda sinptica. Os mediadores qumicos so reconhecidos por receptores especficos da clula seguinte, e a ativao dos receptores na membrana ps-sinptica capaz de converter o sinal qumico em sinal eltrico novamente.
- 42 -
4.4. Integrao dos Sinais
A maioria das clulas nervosas recebe diversas aferncias numa mesma regio, e todo o conjunto de estmulos, sejam despolarizantes ou hiperpolarizantes (potencial excitatrio pssinptico PEPS; potencial inibitrio ps-sinptico PIPS, respectivamente). Esse conjunto de sinais integrado e a corrente eltrica resultante pode, ou no, deflagrar o potencial de ao na zona de disparo.
4.5. LTP
O LTP (long-term potentiation ou potenciao de longa durao) um aumento na durao da transmisso sinptica entre dois neurnios, que resulta na estimulao de ambos de modo sincrnico. Est associado plasticidade sinptica, ou seja, capacidade das sinapses de se modificarem. Este tipo de evento encontrado principalmente no hipocampo, local intimamente relacionado s memrias. De uma maneira mais detalhada, o LTP aumenta a transmisso sinptica, por aumentar a habilidade de dois neurnios, um pr e um ps-sinptico se comunicarem.
- 43 -
REFERNCIAS BIBLIOGRFICAS
Behavior. 5 Ed. Boston. Paramount, 1994. pags. 19-46 Carlson, N. R. - Structures and functions of cells of the Nervous System. In: Physiology Hof, P. R.; Trapp, B. D.; Vellis, J.; Claudio, L.; Colmann D.R.- The cellular components of the Nervous Tissue. In: Zigmund, M. J.; Bloom, F. E.; Landis, S. C.; Roberts J.L.; Fundamental Neuroscience. San Diego. Academic Press., 1999, pags. 41-70. Guyton, A. R.; Hall, J. E.; Tratado de Fisiologia Mdica. 9Ed. Philadelphia Guanabara Koogan, 1997. Hof, P.R.; Trapp, B.D.; Vellis,J.; Claudio, L.; Colman, D.R. - The cellular components of Nervous Tissue. In: Zigmound, M.J.; Bloom, F.E.; Landis, S.C.; Roberts, J.L.- Fundamental Neuroscience. San Diego. Academic Press., 1999, p. 41-70. Kandel, E. R.; Schwartz, J. H.; Jessel, T. M.; Principles of Neural Science. 4Ed. New York. McGrawHill Companies, 2000. Lent, R. Cem bilhes de neurnios. 1Ed. So Paulo. Atheneu 2001. Schatzerberg, A. F.; Nemeroff, C. B. Textbook of Psychopharmacology The American Psychiatric Publishing. 3 Ed, Washington, DC; London, England, 1995, p. 89 103.
- 44 -
INTRODUO PSICOFARMACOLOGIA
1. NEUROTRANSMISSO QUMICA
Quando falamos em neurotransmisso qumica, nos referimos comunicao neuronal (ou entre neurnios e clulas-alvo) exercida por meio de mediadores qumicos, os neurotransmissores. Para que esses transmissores, ou mensageiros primrios, sejam capazes de levar eficientemente a mensagem de um neurnio emissor (ou pr-sinptico) a outro destinatrio (ou ps sinptico), so necessrias a realizao de diversas etapas, como ilustrado na figura 1.
Figura 1: Ilustrao de um terminal sinptico. Aps captao e acmulo de precursores (1), o neurotransmissor sintetizado (2) e armazenado em vesculas (3). O neurotransmissor liberado interage com seu receptor ps-sinptico (4) ou auto-receptores (5), que regulam a liberao do mesmo (feedback). A ao do neurotransmissor terminada por recaptao pelo transportador de membrana localizado no terminal pr-sinptico (6), por difuso (7), recaptao pelas clulas da glia (8) ou ser degradada(9). (Figura adaptada de Fundamental Neuroscience).
1.1. Sntese e Armazenamento do Neurotransmissor
Os diferentes neurotransmissores so sintetizados a partir de precursores que, muitas vezes, so captados pelo neurnio por um processo de transporte ativo (como no caso dos aminocidos tirosina e triptofano). No citoplasma ou em organelas especficas, o precursor passa pela ao de enzimas especficas at a formao do neurotransmissor. Uma vez formado, o transmissor
- 45 -
transportado ativamente para vesculas sinpticas, onde armazenado. Estas etapas esto representadas na figura 1 com os nmeros 1, 2 e 3, respectivamente.
1.2. Liberao do Neurotransmissor para a Fenda Sinptica
Aps a sinalizao de um impulso nervoso, conduzido pelo neurnio at o terminal do axnio, ocorre a despolarizao neuronal. Em seguida ocorre a abertura de canais de clcio dependentes de voltagem, causando o influxo de clcio para o meio citoplasmtico. O clcio faz com que as vesculas com neurotransmissores se fundam membrana pr-sinptica do neurnio, fazendo com que haja a liberao dos neurotransmissores na fenda sinptica por meio de exocitose. Uma vez na fenda sinptica, os neurotransmissores interagem com estruturas proteicas macromoleculares que apresentam um stio de ligao especfico para o neurotransmissor, os receptores, como representado na figura 1 pelo nmero 4. Os receptores podem estar localizados nos neurnios pr-sinpticos, como no caso dos auto receptores, ou no neurnio ps-sinptico.
1.3. Receptores
A ligao do neurotransmissor ao receptor ps-sinptico faz com que este seja ativado, causando alteraes eltricas e/ou metablicas no neurnio ps-sinptico, alterando assim seu funcionamento. Essas alteraes podem ocorrer rapidamente, dentro de milissegundos, ou mais lentamente, dentro de horas ou dias. O tipo de resposta celular causada pela ativao dos receptores vai depender do tipo de receptor. Alm disso, podem ainda haver outros stios de ligao (alostricos) no receptor aos quais outras molculas moduladoras podem se ligar e alterar a interao do transmissor ao seu stio de ligao assim, modulam a mensagem carregada pelo neurotransmissor. Neurotransmissores que no se ligam a receptores ps-sinpticos podem se ligar em receptores pr-sinpticos. Desta forma eles regulam o funcionamento do prprio neurnio prsinptico, por exemplo, sinalizando que no mais necessria a liberao de neurotransmissores na fenda sinptica. Este mecanismo tambm conhecido como feedback (retroalimentao) negativo. Os neurotransmissores que no se ligaram ao receptor ps-sinptico podem tambm ter diferentes destinos, como veremos no subtpico 1.5. Com base na estrutura molecular, na sua localizao e no mecanismo de transduo podemos distinguir quatro superfamlias de receptores (figura 2): 1) Receptores acoplados a canais inicos (ou ionotrpicos): receptores para diversos neurotransmissores que formam canais seletivos de ons. Sua abertura ocorre em decorrncia da ligao com o neurotransmissor, a partir de alteraes conformacionais. Esses receptores so
- 46 -
chamados tambm de canais inicos controlados por ligantes e conduzem seus sinais pelo influxo ou efluxo de ons especficos, tais como de sdio, potssio, clcio ou cloro. Isto leva alterao do potencial de membrana da clula (com hiperpolarizao se o potencial gerado for inibitrio, ou despolarizao se o potencial for excitatrio). Quando o estmulo despolarizante e atinge o limiar de disparo, o potencial de ao deflagrado no neurnio ps-sinptico at a prxima terminao nervosa. A ativao destes receptores leva a uma alterao eltrica rpida (na escala de milissegundos). Os exemplos incluem o receptor nicotnico de acetilcolina, o receptor GABAA, os receptores de glutamato dos tipos NMDA, AMPA e Cainato e o receptor serotonrgico 5-HT3. 2) Receptores acoplados protena G (ou metabotrpicos): recebem este nome, pois so ligados protena G um trmero composto pelas subunidades , e . Uma vez ocorrida, a ligao do neurotransmissor a estes tipos de receptores, a subunidade da protena G se separa, atuando sobre outras enzimas, como fosfolipases e adenilato ciclase (tambm presentes na membrana celular), ou mesmo canais inicos. As fosfolipases e adenilato ciclase, por sua vez, atuam sobre outros elementos intracelulares, produzindo os chamados segundos mensageiros, como AMP cclico, clcio, inositol trifosfato e outros. Os segundos mensageiros se localizam no neurnio ps-sinptico e so responsveis por dar seguimento sinalizao iniciada pelo neurotransmissor gerando aes celulares e efeitos biolgicos. Os metabotrpicos so a maior famlia de receptores no crebro de humanos e incluem receptores para muitos hormnios e transmissores como, por exemplo, o receptor muscarnico de acetilcolina, os receptores adrenrgicos, os receptores dopaminrgicos, os receptores histaminrgicos e os receptores serotonrgicos, com exceo do receptor 5-HT3. 3) Receptores ligados protena quinase: respondem a mediadores proteicos. So diretamente acoplados a protenas quinases (enzimas que catalisam reaes de fosforilao), controlando o funcionamento de outras protenas e enzimas especficas. Uma vez fosforiladas, essas protenas so ativadas e exercem suas funes especficas por um perodo de tempo, at que outras enzimas, as fosfatases, retirem o grupamento fosfato e a protena volte ao estado de menor ativao. Apresenta um domnio extracelular de ligao conectado a um domnio intracelular por uma nica hlice transmembrana. Em muitos casos, o domnio intracelular de natureza enzimtica (com atividade de protena quinase ou de guanilato ciclase). Estes receptores participam das aes de uma ampla variedade de mediadores proteicos, incluindo fatores de crescimento e citocinas, bem como hormnios, como a insulina e a leptina. 4) Receptores nucleares: regulam a transcrio de genes, com consequente sntese de protenas e produo de efeitos celulares. Alguns esto localizados dentro do ncleo, enquanto outros se localizam no citosol e migram para o compartimento nuclear na presena de ligante. Incluem os receptores de hormnios esteroides, do hormnio tireideo e de outros agentes, como o cido retinico e a vitamina D.
- 47 -
Figura 2: Tipos de receptores celulares e suas aes intracelulares. (Figura e tabela retiradas do livro Farmacologia. Rang, H.P.; Dale, M.M.; Ritter, J.M. e Moore, P.K. Editora Elsevier. 5 edio).
1.4. Transduo do Sinal
No caso dos receptores acoplados a canais inicos e daqueles acoplados a enzimas (como a tirosina quinase), a ativao do receptor produz uma ao celular imediata (mobilizao de ons; fosforilao proteica), que pode gerar outras aes posteriores mais lentas. No caso dos receptores metabotrpicos, a ativao gera reaes moleculares at que sua ao metablica seja efetivada no neurnio ps-sinptico. Se, por um lado isso pode levar lentificao da resposta celular, por outro, a cascata de reaes permite a amplificao do sinal pela produo de segundos mensageiros.
- 48 -
1.5. Trmino da Ao do Neurotransmissor na Fenda Sinptica
Para que o processo de neurotransmisso seja eficaz, faz-se tambm necessria a finalizao da ao do neurotransmissor, garantindo, assim, a prontido do sistema para poder responder a um novo estmulo de maneira eficiente. Para tanto, neurotransmissores liberados devem ser rapidamente removidos ou inativados. Os processos de recaptao e degradao consistem nos principais mecanismos de terminao da ao dos neurotransmissores. Na recaptao, o transmissor retirado da fenda sinptica por protenas transportadoras especficas, que levam as molculas do neurotransmissor para o citoplasma da clula prsinptica. Essa recaptao pode ser feita pelo prprio neurnio pr-sinptico ou por clulas da glia (mecanismo extra neuronal). Assim, o neurotransmissor recaptado pode ser reciclado, sendo armazenado novamente nas vesculas para posterior liberao. O processo de degradao do neurotransmissor executado por enzimas presentes na prpria fenda sinptica ou por enzimas intracelulares. Neste caso, os metablitos podem ser reaproveitados na sntese de novas molculas. Alm destes dois mecanismos, pode tambm ocorrer a disperso do neurotransmissor no meio intersticial, permitindo, por exemplo, que este neurotransmissor se ligue a outros receptores de neurnios prximos quele ao qual ele foi liberado.
2. INTERAO FRMACO-RECEPTOR
A interao frmaco-receptor (F-R) obedece lei de ao das massas (F + R F-R) e, portanto, depende primariamente de fatores estereoqumicos que determinam a natureza da interao. A maioria das drogas favorece a combinao com seus receptores atravs de foras de baixa intensidade (ligaes inicas, foras de Van der Waals e pontes de hidrognio), como ocorre nas molculas endgenas.
2.1. Agonistas
So substncias que se ligam ao receptor, ativando-o. Dentre essas substncias podemos citar os neurotransmissores (agonistas endgenos) e alguns psicofrmacos, que mimetizam a ao dos prprios neurotransmissores. Os agonistas podem ser subdivididos em trs tipos: a) agonistas plenos: aqueles capazes de ativar o receptor de maneira total, ou seja, sua ligao no receptor gera um efeito completo;
- 49 -
b) agonistas parciais: apesar de estarem ligados ao receptor, no provocam uma resposta completa, quando comparados aos agonistas plenos (figura 3A); c) agonistas inversos: ativam o receptor, porm causam um efeito inverso ao dos agonistas totais ou parciais. Por exemplo, o frmaco sarmazenil que pertence famlia dos benzodiazepnicos, ao invs de diminuir a ansiedade (como a maioria dos benzodiazepnicos), aumenta a mesma. Logo, sua curva doseefeito seria a de um agonista total ou parcial, porm de forma espelhada.
2.2. Antagonistas
Um antagonista se liga ao receptor sem, no entanto desencadear nenhum tipo de ao. O antagonista pode se ligar ao mesmo stio do agonista ou a um stio diferente, conforme veremos mais a frente. Tais compostos no possuem atividade regulatria intrnseca, logo, no ativam o receptor, mas produzem efeitos por inibio da ao dos agonistas. Antagonismo fisiolgico ocorre quando h interao de duas substncias cujas aes opostas no organismo tendem a anular uma a outra, no atuando no mesmo receptor. Por exemplo, em uma situao de anafilaxia, a histamina atua sobre receptores das clulas pulmonares causando broncoespasmo. Neste caso, recorrente a administrao de adrenalina, que atua inibindo os efeitos da histamina, mesmo agindo em outro receptor. Antagonismo especfico ou farmacolgico ocorre quando agonista e antagonista agem sobre o mesmo receptor. Este tipo de antagonismo pode ocorrer de duas formas: a) antagonismo competitivo geralmente substncias de estrutura anloga, competem por um mesmo stio de ligao no receptor. caracterizado pelo deslocamento da curva doseresposta para a direita, uma vez que necessrio aumento da concentrao do agonista para se promover o mesmo efeito. Pode ser subdividido em antagonismo competitivo reversvel (o aumento da dose do agonista pode levar ao desacoplamento do antagonista do receptor) e antagonismo competitivo irreversvel (o aumento da concentrao do agonista no suficiente para desacoplar o antagonista de seu stio de ligao). Neste caso, a sntese de um novo receptor necessria. b) antagonismo no competitivo geralmente sua ligao ocorre em stios diferentes do utilizado pelo neurotransmissor no receptor, promovendo uma alterao conformacional do mesmo, impedindo o agonista de se ligar ou que o complexo agonista-receptor seja ativado. Por no agir no mesmo stio de ligao, no possvel desfazer o bloqueio aumentando a concentrao do agonista. Nesse caso, a curva dose-resposta em presena do agonista no se desloca paralelamente e sim, muda de inclinao. O grau de bloqueio s depende da dose do antagonista (figura 3B).
- 50 -
Figura 3: (A) Curva dose-resposta para o agonista total (resposta mxima corresponde a 100% do efeito) e para o agonista parcial (resposta mxima no atinge 100% do efeito). (B) Curva de ocupao do receptor pelo agonista na presena do antagonista. Observe que o antagonista competitivo produz deslocamento da curva para a direita, sem diminuir o efeito mximo, enquanto o antagonista no competitivo provoca diminuio do efeito.
Antagonismo qumico Ocorre quando o antagonista reage quimicamente com o agonista, inativando-o. Dentre os exemplos destaca-se o uso de agentes quelantes que se ligam a metais pesados, reduzindo sua toxicidade. Antagonismo farmacocintico: um frmaco interfere na absoro, distribuio, metabolismo ou excreo de um outro. Por exemplo, o antibitico tetraciclina no deve ser administrado juntamente com anticidos (contendo clcio ou magnsio), j que estes complexam com a tetraciclina, impedindo sua absoro.
2.3. Afinidade e Eficcia
Em geral, o aumento da concentrao do agonista produz um aumento do efeito desencadeado pelo receptor. No entanto, nem sempre h uma relao clara e direta entre a quantidade de receptores ocupados pelo agonista e seu efeito. Isto ocorre porque a afinidade pelo receptor diferente para cada frmaco, e tambm porque cada agonista pode estimular o receptor com maior ou menor intensidade (o que chamado de eficcia). Em outras palavras, a afinidade representa a facilidade pela qual o frmaco se liga ao receptor, enquanto eficcia representa a capacidade deste de, uma vez ligado, provocar mudanas no receptor que levem aos seus efeitos.
- 51 -
2.4. Interao Entre Frmacos
Ocorre quando os efeitos de um frmaco so alterados pela presena de outro frmaco, alimento, bebida ou algum agente qumico ambiental. Frmacos que agem sobre o mesmo tipo de receptor, ou sobre receptores que modulam a mesma funo, podem produzir os seguintes efeitos: a) sinergismo de efeito aditivo soma dos efeitos de dois frmacos; b) potenciao ocorre quando o efeito combinado dos dois frmacos maior que a soma dos efeitos isolados.
3. REGULAO DE RECEPTORES
A resposta dos receptores aos seus respectivos neurotransmissores dinmica, e pode ser modulada por diversos fatores. Dentre esses, j foi citada a modulao do efeito do receptor por stios alostricos de ligao e receptores, cujo funcionamento modulado pela fosforilao. Alm disso, dois fenmenos modulam a ativao do receptor: a) dessensibilizao (ou downregulation): aps a ativao continuada dos receptores por agonistas, o efeito subsequente de uma mesma concentrao do agonista diminudo (tolerncia). b) supersensibilidade (ou upregulation): aumento da resposta de um receptor devido continuada inibio do mesmo. Esses processos homeostticos modulam a resposta dos receptores, e podem ser resultantes, em alguns casos, da alterao da afinidade ou do nmero de receptores presentes na membrana celular. Em casos de dessensibilizao, a resposta celular compensatria seria no sentido de diminuir a eficincia no funcionamento do receptor, enquanto que na supersensibilidade a resposta compensatria aumenta a eficincia do receptor.
4. PRINCIPAIS VIAS DE NEUROTRANSMISSO
4.1. Acetilcolina (ACh)
A acetilcolina um neurotransmissor responsvel pela mediao de uma srie de processos que envolvem tanto o sistema nervoso central quanto o perifrico. Sntese (figura 4): A sntese da acetilcolina no SNC ocorre em neurnios prprios e se d pela ao da enzima colina acetil transferase. Esta enzima realiza a transferncia do radical acetil, proveniente de acetil coenzima A (Acetil CoA), para a colina. A colina uma vitamina do
- 52 -
complexo B, derivada de fontes alimentares, e Acetil CoA, produzido a partir de glicose e cidos graxos. Receptores: As funes da ACh so mediadas por dois tipos diferentes de receptores: nicotnicos (ionotrpicos) e muscarnicos (metabotrpicos). Os receptores muscarnicos apresentam tanto aes excitatrias quanto inibitrias e so subdivididos em cinco: M1, M2, M3, M4 e M5. J os receptores nicotnicos tem ao apenas excitatria. No SNC, estes receptores so amplamente distribudos, embora a densidade dos nicotnicos seja menor do que a dos muscarnicos. Degradao: Processo responsvel pela terminao da ao, e dado pela ao de 2 enzimas: Acetilcolinesterase e butirilcolinesterase. A acetilcolinesterase encontrada em neurnios do SNC em geral, e apresenta atividade de degradao especifica para a Ach. A butirilcolinesterase uma enzima principalmente srica e responsvel pela degradao de uma serie de steres, entre eles a Ach, portanto com atividade inespecfica. Pela ao dessas enzimas, a ACh hidrolisada em colina e acetato, sendo a colina novamente captada pela terminao pr-sinptica.
Figura 4: Sntese e degradao da Acetilcolina. Vias neuronais (figura 5): Existem dois grandes grupos de neurnios colinrgicos no encfalo: (1) grupo caudal, localizado no tronco enceflico, composto pelos ncleos tegmentais ltero-dorsal e pednculo pontino, que se projetam para a formao reticular da ponte e do bulbo, para o cerebelo e, principalmente, para o tlamo; e (2) o complexo colinrgico do prosencfalo basal, composto pelos ncleos septal medial, ncleo da banda diagonal, substncia inonimata, rea pr-ptica e ncleo basal de Meynert, que se projetam principalmente para o hipocampo e crtex cerebral. Alm dessas vias, interneurnios colinrgicos curtos so tambm encontrados em algumas regies, como no estriado e no nucleus accumbens. Funes: As principais funes das vias colinrgicas esto relacionadas com viglia, aprendizado e memria. Alm de suas funes no Sistema Nervoso Autnomo, como bradicardia, midrase, e motilidade intestinal.
- 53 -
Figura 6: Julius Axelrod.
Figura 5: Vias da Acetilcolina no crebro. A localizao dos principais grupos de corpos celulares e tratos de fibras mostrada em vermelho. (1- Ncleo basal; 2 - Crtex; 3 - Septo; 4 - Hipocampo; 5 - Substncia Negra; 6 - Tlamo). Adaptado de http://webpages.fc.ul.pt/~rfcruz/relats/reltlb13.html
4.2. Catecolaminas
Por volta da dcada de 50, o mdico e bioqumico, Julius Axelrod (figura 6), ao estudar os efeitos e mecanismos de ao da cafena no sistema nervoso central, realizou descobertas sobre o sistema nervoso simptico e seus componentes, como o hormnio Epinefrina (Adrenalina), e os neurotransmissores Norepinefrina (ou Noradrenalina) e Dopamina. As catecolaminas formam um grupo que tem como seus principais componentes a norepinefrina, epinefrina e dopamina, e so assim denominadas, por possurem em sua estrutura um grupo catecol e uma amina de cadeia lateral. Sntese: ocorre inicialmente no axnio e terminada j dentro das vesculas secretrias da clula. Ela pode ser iniciada diretamente pelo aminocido tirosina, ou pelo aminocido fenilalanina, que convertido em tirosina durante a sntese. A rota de sntese da norepinefrina, epinefrina e dopamina a mesma, porm, dependendo da etapa que a rota interrompida, originar uma determinada catecolamina. Quem determina em qual parte da rota a sntese ser interrompida a prpria clula, por exemplo, os neurnios dopaminrgicos no produzem a enzima dopamina hidroxilase, portanto no possvel transformar a dopamina em norepinefrina . Na figura 7 podemos observar a rota passo a passo.
- 54 -
Figura 7: Biossntese das principais catecolaminas. 1- Fenilalanina hidroxilase, 2- Tirosina hidroxilase, 3- Dopa descarboxilase, 4- Dopamina hidroxilase e 5- Feniletanolamina N-metil-transferase. Adaptado de http://www.qmc.ufsc.br/qmcweb/artigos/adrenalina.html Degradao: ocorre de trs maneiras 1) recaptao pela prpria terminao nervosa por transporte ativo, 2) difuso para tecidos adjacentes e, 3) degradao por enzimas, como a Monoamino oxidase (MAO), encontrada nas terminaos nervosas, e a Catecol-ortoaminotransferase (COMT), encontrada em vrios tecidos. Seus metablitos, como por exemplo, o cido dihidroxifenilactico e o cido homovanlico, no caso da dopamina; o cido vanilmandlico, principalmente na periferia, e o 3-metoxi, 4-hidroxifenilglicol, principalmente no crebro, para a noradrenalina, so todos excretados na urina e podem ser analisados e quantificados.
4.2.1. Dopamina (DA)
Receptores: Os receptores da dopamina so divididos em duas famlias principais: a famlia D1, que inclui os subtipos D1 e D5, e a famlia D2, composta pelos subtipos D2, D3 e D4. Todos estes subtipos so metabotrpicos, sendo que a famlia D1 est associada ativao da adenilato ciclase, enquanto a famlia D2, inibio da mesma. Vias neuronais (figura 8): A dopamina apresenta vias bem definidas no SNC. As principais so: Mesolmbica neurnios localizados no tegmento mesenceflico (rea tegmental ventral, VTA) projetam-se para as reas lmbicas (amgdala, ncleo accumbens, septo, tubrculo olfatrio e crtex piriforme).
- 55 -
Mesocortical neurnios da VTA projetam-se para os crtices pr-frontal, medial, do cngulo e entorrinal. Nigro-estriatal neurnios da substncia negra projetam-se para o estriado. Tbero-infundibular projees dos ncleos arqueados e periventricular do hipotlamo para a eminncia mediana e lobo intermedirio da hipfise. Funes: As principais funes da dopamina relacionam-se com as suas vias neurais: controle neuroendcrino, pela via tbero-infundibular; controle motor, pela via nigro-estriatal; e efeitos comportamentais e cognitivos, pelas vias mesolmbica e mesocortical.
Figura 8: Vias dopaminrgicas no crebro. Adaptado de Chapman and Hall, 3rd edition.
4.2.2. Norepinefrina (NA)
A maioria dos corpos celulares dos neurnios noradrenrgicos no crebro encontram-se no locus coeruleus (ou locus ceruleus), uma regio localizada no tronco enceflico.
- 56 -
Receptores: Existem duas classes de receptores metabotrpicos aos quais a NA pode se ligar, os receptores do tipo , que se dividem nos subtipos 1, 2 e 3, e receptores do tipo , que se dividem nos subtipos, 1, 2 e 3. Todos os subtipos de receptores noradrenrgicos so receptores ps-sinpticos, exceto pelo receptor 2, que pode estar localizado pr-sinapticamente e agir como auto receptor. Vias neuronais e funes (figura 9): A NA possui vias ascendentes e descendentes. As vias descendentes de transmisso noradrenrgicas esto envolvidas no controle autonmico e visceral (como controle da frequncia cardaca, presso sangunea e esvaziamento da bexiga), e controle da dor. As vias ascendentes projetam-se para diversas reas do crebro, como crtex pr-frontal, hipotlamo, amgdala, e cerebelo, estando envolvidas na modulao do humor, cognio, fadiga, agitao psicomotora, movimentos motores e viglia.
Figura 9: Vias da Noradrenalina no crebro. A localizao dos principais grupos de corpos celulares e tratos de fibras so mostrados em verde. Adaptado de Page, Curtis, Sutter et al, 1997.
4.3. Serotonina (5-HT ou 5-hidroxitriptamina)
A serotonina foi descoberta em 1947 como um substrato responsvel pela vasoconstrio. O nome serotonina um acrnimo composto por serum (soro) e tonin (tnus - propriedade vasotnica). A serotonina pertence classe das aminas biognicas e sua biossntese assemelha-se a das catecolaminas.
- 57 -
Os neurnios produtores de serotonina se encontram no mesencfalo, especificamente nos ncleos da rafe da linha mdia da ponte e no bulbo. Tais neurnios projetam-se especialmente para o crtex, alguns componentes do sistema lmbico (como amgdala e hipocampo dorsal), hipotlamo, e medula espinhal, conforme mostrado na figura 10.
Figura 10: Vias neurais do Sistema Serotoninrgico. Adaptado de Neurotransmitters and drugs, 3rd edition.
Sntese: ocorre a partir do aminocido triptofano e ao da enzima triptofano -hidroxilase, que considerado o fator limitante para a disponibilidade desse neurotransmissor no sistema nervoso. O 5-hidroxitriptofano resultante desta reao convertido a serotonina pela ao da descarboxilase de aminocidos aromticos. Recaptao e degradao: depois de liberada na fenda sinptica, a serotonina pode ser recaptada por transportadores especficos, localizados no neurnio pr-sinptico. Ela pode ser reencapsulada em uma vescula sinptica para novamente ser liberada, ou degradada ao seu metablito, o cido 5-hidroxiindolactico (5-HIAA), pelas enzimas MAO (monoamina oxidase) e aldedo desidrogenase. A excreo de 5-HIAA pela urina proporciona uma medida da renovao de 5-HT. O esquema representando as etapas da sntese e degradao da serotonina pode ser visto na figura 11.
- 58 -
Figura 11: Etapas de sntese (A) e degradao (B) da serotonina (adaptado de Halbach, 2006). Os receptores serotoninrgicos podem ser agrupados em trs diferentes estruturas moleculares: os transportadores, os canais inicos e os acoplados protena G. A classificao dos subtipos dos receptores baseada na sua estrutura e propriedades farmacolgicas, conforme mostrado na tabela abaixo. Subtipos de receptores de serotonina Receptores acoplados protena G Famlia de receptores de 5-HT1 Famlia de receptores de 5-HT2 Outros receptores Canais inicos ligante-dependente Transportadores Subtipos 5-HT1A, 5-HT1B, 5-HT1D, 5-HT1E, 5-HT1F 5-HT2A, 5-HT2B, 5-HT2C 5-HT4S, 5-HT47, 5-HT5A, 5-HT5B, 5-HT6, 5-HT7 5-HT3 Captadores de serotonina
Tabela 1: Classificao dos subtipos dos receptores de 5-HT (adaptado de Halbach, 2006).
Funes: devido a grande diversidade de receptores e diferentes vias de transduo de sinais ativadas, o sistema serotoninrgico responsvel por diversas funes no sistema nervoso central. Os nveis de serotonina influenciam, por exemplo, processos relacionados com a aprendizagem e memria, comportamento sexual e alimentar. Ela tem participao em processos de dimorfismo sexual uma vez que se pode observar diferenas nos nveis de expresso de RNAm de receptores 5-HT1A entre ratos machos e fmeas. A serotonina tambm est envolvida no controle do humor e da emoo, como j foi visto que camundongos knockout para 5-HT1B so mais agressivos. Clinicamente, as alteraes nos nveis de serotonina esto relacionadas com transtornos de ansiedade, afetivos, alimentares e de sono.
- 59 -
4.4. Aminocidos Como Neurotransmissores
Os aminocidos possuem sua funo estrutural e nutricional bem caracterizada, mas alguns deles desempenham funes diferentes a estas, como papel na neurotransmisso. Temos como principais exemplos de aminocidos neurotransmissores o glutamato e cido aminobutrico (GABA), excitatrio e inibitrio, respectivamente. A descoberta e caracterizao de aminocidos como neurotransmissores iniciaram-se em 1950, mas sua aceitao ocorreu somente na dcada de 1970. Um fator importante que auxiliou na aceitao do papel destes compostos foi a descoberta de antagonistas dos aminocidos excitatrios.
4.4.1. Aminocidos Excitatrios
O glutamato o principal neurotransmissor excitatrio, presente em neurnios amplamente distribudos pelo SNC. Possui papel metablico e de neurotransmissor, conforme a ao das enzimas transaminases, que catalisam a interconverso de glutamato e -cetoglutarato. Sua sntese pode ocorrer por duas vias: atravs da glicose, por intermdio do Ciclo de Krebs, ou da glutamina, sintetizada pelas clulas gliais e internalizada pelos neurnios, onde convertida a glutamato (figura 12). A primeira via responsvel por aproximadamente 20% do glutamato, e a segunda, pelo restante (80%).
Figura 12: A: Frmula estrutural do glutamato. B: Metabolismo dos aminocidos neurotransmissores. Os neurotransmissores esto nas caixas pretas e as enzimas em caixas claras (adaptado de Rang e Dale, 6 edio). Depois de sintetizado, o glutamato internalizado e armazenado em vesculas. Este mecanismo protetor impede a degradao do neurotransmissor e realizado pelo transportador
- 60 -
vesicular de glutamato (VGlut). Em resposta ao potencial de ao e consequente influxo de Ca2+, o glutamato ser liberado na fenda sinptica por exocitose (figura 13), podendo interagir com seus receptores ou ser recaptado. Recaptao e degradao: a recaptao pode ser realizada pelo prprio neurnio, mas ocorre principalmente por clulas gliais (astrcitos), sendo a principal forma de controle da excitao do glutamato. Este controle necessrio, visto que excesso deste neurotransmissor na fenda causa morte neuronal (excitotoxicidade). Depois de recaptado, o glutamato pode ser convertido a glutamina, -ceto glutarato, ou ainda no neurotransmissor inibitrio GABA (ver figura 13). Os astrcitos adjacentes convertem o glutamato glutamina, reciclando-o. Dessa forma, os astrcitos participam tanto da sntese como da captao da fenda sinptica.
Figura 13: Transporte de glutamato (Glu) e glutamina (Gln) pelos neurnios e astrcitos. O glutamato liberado internalizado em parte pelos neurnios e em parte pelos astrcitos, que convertem a maior parte dele em glutamina. A glutamina liberada por intermdio do transportador GlnT, e os neurnios a captam e sintetizam glutamato. VGlut, transportador de glutamato, TAAE, transportador de aminocido excitatrio (adaptado de Rang e Dale, 6 edio).
Ao: Atualmente so identificados quatro tipos de receptores, sendo eles divididos em ionotrpicos e metabotrpicos. Os metabotrpicos (mGluR) so receptores acoplados protena G, divididos em 3 famlias (classe I, II e III). So encontrados tanto no pr e ps sinptico, tendo papel modulador.
- 61 -
Os receptores ionotrpicos so classificados em: AMPA, cainato e NMDA. O receptor de cainato foi denominado a partir de seu agonsta especfico, o cido canico; tem distribuio limitada e possvel ao no potencial excitatrio ps sinptico (PEPS). O receptor AMPA consiste em combinaes de subunidades GluR1 a R4. Sua ativao exige a presena de quatro molculas de glutamato e sua permeabilidade varia de acordo com as subunidades constituintes, sendo principalmente ao Na+. Estes receptores so responsveis pela transmisso excitatria rpida do SNC. O receptor NMDA possui caractersticas especiais: altamente permevel ao Ca 2+ (bem como a outros ctions como o Na+), tem seu poro bloqueado pelo Mg2+, e possui uma ativao complexa. Sua ativao exige a ligao da glicina e de glutamato associada a uma despolarizao prvia da membrana. A despolarizao prvia causar mudanas conformacionais no receptor, liberando o bloqueio do canal pelo Mg 2+. Desempenham importante papel nas alteraes adaptativas de longo prazo (plasticidade sinptica) e devido a sua complexa ativao, participam da transmisso excitatria lenta. Receptores de glutamato so mais abundantes no crtex, ncleos da base e nas vias sensitivas. Os receptores AMPA e NMDA so geralmente encontrados co-localizados, isto porque a despolarizao prvia da membrana, exigida pelo NMDA, ocorre em decorrncia da ativao do AMPA. Funo: alm de sua funo excitatria geral participa da estimulao de vias especficas, como processos de plasticidade sinptica. Plasticidade sinptica um termo geral para designar alteraes de longo prazo nas sinapses, sejam elas decorrentes de alteraes fisiolgicas (aprendizagem e memria) ou patolgicas (epilepsia e dependncia). Um componente importante para a plasticidade neuronal a potenciao de longo prazo, no qual os receptores de glutamato desempenham papel central.
4.4.2. Gaba
O cido amino butrico (GABA) um dos principais neurotransmissores do SNC, e predominantemente inibitrio. Apresenta ao excitatria no perodo embrionrio e ps-natal imediato. Os neurnios GABArgicos so fundamentalmente interneurnios, cuja funo regular a excitabilidade neuronal. So distribudos por toda a substncia cinzenta, mas com abundncia no sistema nigroestriatal. Sntese: ocorre a partir do glutamato e da ao da enzima descarboxilase do cido glutmico (DAG). Essa enzima responsvel pela descarboxilo do glutamato e encontrada apenas nos neurnios GABrgicos.
- 62 -
A regulao ocorre de duas maneiras: em casos de excesso de GABA a enzima passa para sua forma inativa, j em situaes de intensa ativao neuronal, h maior utilizao de ATP, levando a maior formao de fosfato inorgnico, que age retornando DAG a seu estado ativo. Aps sintetizado, o GABA ser internalizado em vesculas atravs de transporte ativo. Da mesma forma que para os demais neurotransmissores, a sua liberao depende do influxo de Ca2+ por consequncia do potencial de ao. Recaptao e degradao: aps sua liberao, o GABA pode ser recaptado ou interagir com seus receptores. A recaptao o mais importante mecanismo para controlar a concentrao do neurotransmissor na fenda sinptica. Estes transportadores (GATs) esto presentes tanto nos neurnios como em tecidos adjacentes e internalizam o GABA atravs da troca ativa com ons de Cl- e Na+. Uma vez recaptado, o GABA sofre degradao, catalisada pela GABA transaminase, na qual o grupo amino transferido para o -cetoglutarato, produzindo glutamato (figura 14). Ao: A interao do GABA com seus receptores resulta em um influxo de nions, causando hiperpolarizao da membrana do neurnio, tornando-o menos responsivo. O GABA possui trs tipos de receptores: dois deles ionotrpicos, GABAA e GABAC, e um metabotrpico, GABAB. Os receptores GABAA sofrem modulao alostrica por diferentes compostos, como os barbitricos, benzodiazepnicos e lcool. Estes compostos possuem ao inibitria para o SNC, pois auxiliam o mecanismo de abertura destes canais inicos. Os receptores GABAB so receptores acoplados protena G presentes tanto na membrana pr como ps-sinptica: na pr sinptica quando ativados resultam no controle da liberao de GABA, enquanto que a ativao dos ps sinpticos resulta em inibio neuronal. Os receptores GABAC so encontrados em neurnios da retina e da hipfise. Pouco se sabe a respeito de suas funes, porm sua ativao permite o influxo de carboxilas (HCO 3-).
Figura 14: A: Formula estrutural do GABA. B: Metabolismo do GABA. Os neurotransmissores esto nas caixas pretas e as enzimas em caixas cinza (adaptado de Rang e Dale, 6 edio).
- 63 -
Funo: o GABA apresenta papel importante na regulao da atividade neuronal, sendo assim, todo seu sistema pode ser visto com alvo teraputico em doenas de hiperexcitabilidade neuronal, como epilepsia.
4.4.3. Glicina
A glicina o principal neurotransmissor inibitrio do tronco enceflico e da medula espinhal, alm disso, desempenha papel facilitador da ativao dos receptores glutamatrgicos NMDA. Sntese: ocorre a partir do aminocido serina pela ao da enzima serina hidroximetil transferase. Essa reao tem como cofator a vitamina B6. O receptor de glicina assemelha-se ao receptor GABAA, sendo um canal multimrico de cloro ativado por ligante.
4.5. Sistema Endocanabinide
Apesar de amplamente utilizada na medicina popular h sculos, a Cannabis sativa (maconha) s teve seu principal composto ativo, o 9-tetrahidrocanabinol, isolado em 1965. Aps essa descoberta comeou-se a estudar os efeitos biomoleculares deste composto e foi identificada a sua capacidade de ativar especificamente determinados receptores. Tendo em vista que uma substncia exgena possui a capacidade de ativar receptores celulares, chegouse a concluso da existncia de um sistema endgeno com as mesmas propriedades. Este sistema foi batizado de endocanabinide. O isolamento do 9 tetrahidrocanabinol deu incio a uma grande corrida no estudo deste composto e de seus efeitos, contudo, os principais endocanabinides e seus receptores especficos s foram identificados trs dcadas depois. Em 1990, o primeiro receptor canabinide foi identificado e denominado de CB1, j em 1993, um segundo receptor foi identificado e denominado de CB2. Esses receptores so do tipo metabotrpicos e so expressos no sistema nervoso central (CB1/CB2) e no sistema imune (CB2). O primeiro endocanabinide identificado foi a araquinoetanolamida ou anandamida (AEA), em 1992. Em seguida foi identificado outro endocanabinide, o 2-araquidonil glicerol (2AG). Atualmente j foram identificados outros endocanabinides como o ter de noladina, a virodamina e a N-araquidonil dopamina (NADA) entre outros, no entanto, a maioria das pesquisas cientficas concentra-se sobre a AEA e o 2-AG. Alm disso, possveis candidatos a CB3, como o G protein-coupled receptor 55 (GPR 55), encontram-se em fase de estudo. Os endocanabinides atuam no sistema nervoso central como mensageiros retrgrados, ou seja, so liberados pelos neurnios ps-sinpticos e atuam em receptores presentes nos prsinpticos. Esses neuromoduladores so sintetizados por demanda e no so armazenados em
- 64 -
vesculas sinpticas (tanto esse fator quanto sua sinalizao retrgrada os diferenciam dos neurotransmissores clssicos, por isso no so classificados como tal). Sntese: Ativao de receptores metabotrpicos especficos e influxo de Ca +2 funcionam como disparadores da atividade de enzimas como a N-acetil transferase e fosfolipase D, responsveis pela converso de fosfatidiletanolamidas em AEA, e da DAG lipase, responsvel pela converso do diacilglicerol em 2-AG. Os endocanabinides atingem a fenda sinptica atravs da atividade de protenas transportadores presentes na membrana celular. Ao: Agem Tanto os receptores CB1 quanto CB2 ativam preferencialmente protena G i/0, esta por sua vez: reduz a atividade da adenilato ciclase, consequentemente reduzem os nveis intracelulares de AMPc; favorecendo a abertura de canais de K+ e fechamento de canais de Ca+2, levando a hiperpolarizao da membrana. Aps exercerem sua atividade, os endocanabinides so recaptados pelo neurnio pssinptico e so degradados por enzimas intracelulares como a FAAH (AEA) e MAG lipase (2-AG). Funo: Os endocanabinides esto envolvidos com diversos mecanismos fisiolgicos, tanto centrais quanto perifricos. Alguns exemplos do envolvimento do sistema endocanabinide com o controle fisiolgico corporal so: Controle da espermatognese em homens Regulao da homeostase Modulao de comportamentos motivados como alimentao e reproduo Controle da ansiedade e da dor
4.6. xido Ntrico (NO)
De maneira independente, a fisiologia circulatria e a neural foram responsveis pelos primeiros estudos a respeito do xido ntrico como sinalizador. Destacam-se dois estudos realizados nos anos 70 e comeo dos anos 80. O primeiro estudo observou que a sinalizao por glutamato favorecia o aumento de guanina monofosfato cclica (GMPc) dependente de Ca +2, o que sugeria a presena de um sinalizador intermedirio desconhecido, j que o glutamato por si no capaz de causar esse efeito. O segundo estudo identificou a presena de uma substncia no endotlio que era capaz de causar um relaxamento da musculatura aps exposio acetilcolina, essa substncia foi chamada de fator de relaxamento derivado do endotlio (FRDE) . Estudos seguintes de ambas as reas observaram que esses efeitos eram: dependentes da ativao da guanilato ciclase e da presena de L-arginina, estes efeitos eram mimetizados com a exposio a agentes liberadores de NO, como o nitroprussiato. Apenas no final dos anos 80 tanto o FRDE quanto o sinalizador desconhecido foram identificados como sendo o NO e deu-se incio a identificao da via de sntese e mecanismos de ao de sinalizador.
- 65 -
Assim como outras substncias sinalizadoras do SNC, o NO no classificado como neurotransmissor, isso porque ele: no armazenado em vesculas sinpticas, no liberado por exocitose e no apresenta receptores extracelulares especficos. Sntese: o aumento intracelular de Ca+2 o primeiro passo na sntese do NO. Esse efeito pode ser induzido pela ao de diferentes neurotransmissores como glutamato, acetilcolina, substncia P, histamina e bradicinina. A calmodulina e o Ca +2 iro formar um complexo responsvel pela ativao da NO-sintase (NOS). Esta enzima encontrada em neurnios (nNOS), no endotlio (eNOS) e uma terceira forma induzvel (iNOS). Seu mecanismo de ao consiste na converso da L-arginina em NO e citrulina, para tal ela utiliza o NADPH e a tetrahidrobpterina como co-fatores. Aps sua sntese, o NO difunde-se para o meio externo e assim capaz de atingir outras clulas adjacentes. Por ser um gs, sua difuso facilitada para o meio intracelular, onde ativa a guanilato ciclase solvel. Esta enzima encontra-se expressa em diversos tecidos como o crebro, pulmo e endotlio. Sua ao consiste em converter o trifosfato de guanina (GTP) em monofosfato de guanina cclico (cGMP). O cGMP atua como segundo mensageiro intracelular e ativa a protena cinase G (PKG), diferentes tipos de fosfodiesterases e canais de Na+ e Ca+2. O NO inativado atravs da sua combinao com o radical heme da hemoglobina ou por sua oxidao a nitrito e nitrato sendo excretado na urina. Ao: Os principais mecanismos que envolvem o NO esto relacionados com o sistema circulatrio e SNC, como: Neurognese e neuroproteo Formao de memria Controle de comportamentos motivados como alimentao e reproduo Relaxamento da musculatura lisa vascular
4.7. Neuropeptdeos
Neuropeptdeos
so
molculas
de
origem
proteica
apresentam
funes
de
neurotransmisso e modulao. Esto envolvidos em quase todos os processos no sistema nervoso, como analgesia (ex: endorfinas), aprendizado (ex: angiotensina IV), controle de presso arterial (ex: angiotensina II), reproduo sexual (ex: ocitocina), controle de funo renal (ex: vasopressina), entre outros. Sntese: Neuropeptdeos so sintetizados a partir de molculas proteicas chamadas de propeptdeos que, ao sofrerem ao de peptidases especficas, liberam os neuropeptdeos correspondentes. Alguns propeptideos liberam mais de um neuropeptideo, por exemplo, a proopiomelanocortina (POMC) que origina diversos neuropeptideos, como -endorfina, MSH, ACTH, entre outros (figura 15).
- 66 -
Figura 15: Formao de neuropeptdeos a partir de POMC, que ao sofrer ao das enzimas prohormnio convertase 1(PC 1) e prohormnio convertase 2 (PC 2) libera neuropeptdeos como ACTH, -endorfina, -MSH, entre outros (adaptado de Goodman e Gilman, 11 edio).
Funo:
Ligam-se
seus
receptores
especficos,
que
em
grande
maioria
so
metabotrpicos, desencadeando as mais diversas funes no SNC. Comunicao celular: Neuropeptdeos podem exercer sua funo utilizando diferentes tipos de comunicao celular (figura 16): a) Autcrino ao na mesma clula em que produzido. b) Parcrino (ou mediadores locais) ao na clula em que produzido e em clulas adjacentes. c) Endcrino Sintetizados em clulas distantes de seus alvos de ao, necessitam de transporte d) Sinptica Atuao como neurotransmissor, desencadeando respostas eltricas no neurnio e) Neuro-hormonal Ao como mediadores em clulas nervosas. Assim como na comunicao endcrina, sua sntese realizada distante da clula alvo.
- 67 -
Figura 16: Comunicao bioqumica de exercida por peptdeos (adaptado de Peptides: Chemistry and Biology, Norbert Sewald and Hans-Dieter Jakuble, 2002).
Apesar de atuar, em alguns casos, como neurotransmissores, os neuropeptdeos apresentam grandes diferenas quando comparados aos clssicos (tabela 2).
Neurotransmissor clssico Pequenas, solveis em gua, Tipo de molcula compostas por grupos amina e carboxila Precursores provenientes da dieta, Sntese enzimas terminal citoplasma Estoque Em pequenas vesculas Enzimas de sntese e vesculas Transporte de estoque so transportadas pelo axnio sintetizados especficas, nervoso ou por no no
Neuropeptdeos Molculas compostas aminocidos Transcrio e traduo de genes So de prpeptdeos. aps originados grandes, por
ativao das peptidases. Ocorre no corpo celular Em grandes vesculas
Somente as vesculas so transportadas pelo axnio
- 68 -
Por Liberao
exocitose em
do
terminal regies
nervoso
Por exocitose, mas no em zonas ativas
especializadas (zonas ativas) Ao Parcrina No Metabolismo no interior do Inativao neurnio ou na fenda sinptica, ou inativao via recaptao
Sim Catabolismo peptidases, recaptao reutilizao por sem ou
Tabela 2: Comparao entre neurotransmissores e neuropeptdios (Belzung Et AL, 2005)
4.8. Sistema de Opiides Endgenos
A nomenclatura do sistema opiide proveniente das substancia derivadas do pio. Estas substncias j foram denominadas de narcticos, hipnoanalgsicos, opiceos, e mais recentemente opiides. O termo opiide foi proposto para designar as drogas com ao semelhante da morfina, porm com estrutura qumica diferente. No entanto, o conceito de opiide evoluiu e passou a incluir todas as substncias naturais, semi-sintticas ou sintticas que reagem com os receptores opiides, seja como agonista ou como antagonista Dentre os opiides naturais temos os endgenos, que se apresentam em sistemas de neuropeptdeos. Possuem funo na modulao da nocicepo (dor e analgesia), alm de estar envolvido no controle de temperatura e reflexos, como a tosse. Os principais opiides endgenos so: -endorfina, encefalinas e dinorfinas. Sntese: - -endorfina: possui como precursor a pr-opiomelanocortina (POMC), que quando processado por proteases da classe pr-hormnio convertase (PCs), d origem ao ACTH (hormnio adrenocorticotrfico), endorfina. - Encefalinas: met e leu-encefalinas so originadas da pr-encefalina, quando processada pelas PCs. - Dinorfinas: Dinorfinas A e B so originadas da pr-dinorfina, quando processada pelas PCs. Recentemente a formao de dinorfinas a partir de pr-dinorfina, tambm foi relacionada com o processamento desta por catepsina L. Receptores: So receptores acoplados protena G e classificados em quatro categorias: (mu), - e -MSH (hormnio estimulante de melancitos) e -
- 69 -
(delta), k (kappa) e ORL1 (opioid-like receptor). a) (delta): subdivididos em 1 e 2, esto envolvidos principalmente em analgesia medular, depresso respiratria e constipao intestinal; b) k (kappa): subdivididos em k1, k2 e k3, esto envolvidos principalmente com disforia e sedao. c) (mu): subdivididos em 1, 2 e 3, esto envolvidos principalmente euforia, dependncia fsica, analgesia supramedular e depresso respiratria; d) ORL1 (opioid-like receptor): este receptor foi descoberto em meados da dcada de 90 e ainda no bem definido, no entanto est relacionado com propriedades ansiolticas e analgsicas. Vias neurais: se originam do hipotlamo e se projetam para diversas reas do sistema nervoso central, como estruturas do sistema lmbico (rea tegmentar ventral e ncleo accumbens). (mu) Analgesia supramedular Analgesia medular Analgesia perifrica Depresso respiratria Miose Euforia Sedao Constipao intestinal Dependncia fsica Disforia Reteno urinaria Prurido Nuseas e vmitos +++ ++ ++ +++ ++ +++ ++ ++ +++ + + (delta) ++ ++ ++ + + k (kappa) + + + ++ + + +++ -
Tabela 3: Efeitos dos receptores opiides (adaptado de Goodman and Gilman, 11 edio)
- 70 -
Trmino da ao sinptica: ocorre atravs da degradao por enzimas proteolticas especficas. - -endorfina: Metabolizada principalmente por catepsina D e E - Leu-encefalina: Metabolizada principalmente por neprilisina, carboxipeptidase A6 e Cistinil aminopeptidase - Met-encefalina: Metabolizada pelas encefalinases (ex: Enzima conversora de angiotensina, Neprilisina, entre outras). - Dinorfina A: Metabolizada principalmente por neurolisina. - Dinorfina B: Metabolizada principalmente por neprilisina e pela Enzima conversora de angiotensina tipo 2. Funes: - -endorfina: apresenta afinidade principalmente pelo receptor 1, mesmo stio de ligao de morfina, sendo assim, relacionado nocicepo, analgesia e bem-estar. Alm disso, a -endorfina apresenta funo no comportamento sexual e apetite. - Met-encefalina: apresenta maior afinidade por receptores do tipo e , e funo analgsica, antidepressiva e papel no sistema imunolgico. - Leu-encefalina: apresenta maior afinidade por receptores do tipo , e funo gonadal - Dinorfina A: apresenta maior afinidade por receptores do tipo k e , e funes relacionadas a estresse, depresso, ciclo circadiano e regulao da temperatura. - Dinorfina B: apresenta maior afinidade por receptores do tipo k e baixa afinidade por e . Suas funes so muito semelhantes dinorfina A. (mu) (delta) k (kappa) -endorfina Leu-encefalina Met-encefalina Dinorfina A Dinorfina B +++ ++ ++ ++ + +++ +++ +++ + +++ +++
Tabela 4: Afinidade das endorfinas pelos seus receptores (adaptado de Goodman e Gilman, 11 edio)
- 71 -
REFERNCIAS BIBLIOGRFICAS:
Belzung, C., I. Yalcin, et al. Neuropeptides in psychiatric diseases: an overview with a particular focus on depression and anxiety disorders. CNS Neurol Disord Drug Targets, v.5, n.2, Apr, p.135-45. 2006. Deguchi T, Yoshioka M (1982) l-Arginine identified as an endogenous activator for soluble guanylate cyclase from neuroblastoma cells J Biol Chem 257(17):1014751 Devane WA, Hanus L, Breuer A, Pertwee RG, Stevenson LA, Griffin G, Gibson D, Mandelbaum A, Etinger A, Mechoulam R (1992) Isolation and structure of a brain constituent that binds to the cannabinoid receptor Science 258(5090):1946-9 Duarte, DF. Uma breve histria do pio e dos opiides. Rev. Bras. Anestesiol., Campinas, v. 55, n. 1, Feb. 2005 . Available from <http://www.scielo.br/scielo.php?script=sci_arttext&pid=S003470942005000100015&lng=en&nrm=iso>. access on 26 Nov. 2012. http://dx.doi.org/10.1590/S003470942005000100015. Ferrendelli JA, Chang MM, Kinscherf DA (1974) Elevation of cyclic GMP levels in central nervous system by excitatory and inhibitory aminoacids J Neurochem 22:53554. Ferrendelli JA, Rubin EH, Kinscherf DA (1976) Influence of divalent cations on regulation of cyclic GMP and cyclic AMP levels in brain tissue J Neurochem 26:741748 Furchgott RF, Zawadzki JV (1980) The obligatory role of endothelial cells in the relaxation of arterial smooth muscle by acetylcholine Nature 288(5789):373-6 Garthwaite J, Charles SL, Chess Williams R (1988) Endothelium-derived relaxing factor release on activation of NMDA receptors suggests role as intercellular messenger in the brain Nature 336:38588 Garthwaite J, Garthwaite G (1987) Cellular origins of cyclic GMP responses to excitatory amino acid receptor agonists in rat cerebellum in vitro J Neurochem 48:2939 Goodman and Gilmans The pharmaceutical basis of therapeutics, 11 edition, 2005. Ignarro LJ, Buga GM, Wood KS, Byrns RE, Chaudhuri G (1987) Endothelium-derived relaxing factor produced and released from artery and vein is nitric oxide Proc Natl Acad Sci USA 84:9265-69. Kruk ZL and Pycock CJ. Dopamine. In: Neurotransmitters and drugs, 3rd edition. London, England: Chapman and Hall, 1993:87115. Matsuda LA, Lolait SJ, Brownstein MJ, Young AC, Bonner TI (1990) Structure of a cannabinoid receptor and functional expression of the cloned cDNA Nature 346(6284):561-4 Mechoulam R, Ben-Shabat S, Hanus L, Ligumsky M, Kaminski NE, Schatz AR, Gopher A, Almog S, Martin BR, Compton DR (1995) Identification of an endogenous 2monoglyceride, present in canine gut, that binds to cannabinoid receptors Biochem Pharmacol 50(1):83-90 Mechoulam R, Gaoni Y (1965) Hashish. IV. The isolation and structure of cannabinolic, cannabidiolic and cannabigerolic acids Tetrahedron. 21(5):1223-9 MEROPS database - http://merops.sanger.ac.uk/
- 72 -
Munro S, Thomas KL, Abu-Shaar M (1993) Molecular characterization of a peripheral receptor for cannabinoids Nature 365(6441):61-5 Page, Curtis, Sutter et al. Drugs in health and disease. In: Integrated Pharmacology. London, UK: Mosby International, 1997: 92151. Ryberg E, Larsson N, Sjgren S, Hjorth S, Hermansson NO, Leonova J, Elebring T,Nilsson K, Drmota T, Greasley PJ (2007) The orphan receptor GPR55 is a novel cannabinoid receptor Br J Pharmacol 152(7):1092-101 Rang, H.P.; Dale, M.M.; Ritter, J.M. e Moore, P.K. Farmacologia. Editora Elsevier. 5 edio. Rang, H.P.; Dale, M.M.; Ritter, J.M. e Moore, P.K. Farmacologia. Editora Elsevier. 6 edio. Sewald, N; Jakuble, HD. Peptides: Chemistry and Biology, online edition, 2002 Squire, Larry R.; Berg, Darwin; Bloom, Floyd E.; du Lac, Sascha; Ghosh, Anirvan; Spitzer, Nicholas C. Fundamental Neuroscience. Academic Press. 3 rd Edition. Von Bohlen und Halbach, Oliver; Dermietzel, Rolf. Neurotransmitters and Neuromodulators Handbook of Receptors and Biological Effects. Wiley-VCH, Weinheim, 2006.
- 73 -
1. MODELOS EXPERIMENTAIS: INTRODUO 1.1. O Conceito de Modelo
A classificao de novos conhecimentos um dos problemas mais antigos na cincia. Por esta razo, uma das prioridades da cincia tem sido a busca por modelos experimentais, a fim de produzir conhecimento de maneira mais padronizada (Ferreira & Ferreira, 2003). Embora a discusso envolvida na conceituao filosfica de modelos seja muito ampla e fuja do objetivo deste texto, algumas consideraes sero feitas a seguir. Historicamente, o termo modulus (do latim, derivado de modus, que significa pequena medida) usado desde 23 A.C. at o sculo XVIII. Do sculo XI ao XIV, o termo utilizado era padro ou molde; do sculo XIV at o XVI, medida e depois modelo (Ferreira & Ferreira, 2003). De acordo com Morato (1993), o termo modelo usado para descrever uma elaborao terica relacionando dois ou mais elementos da Natureza, de tal modo que a descrio dessa relao sirva de base para a compreenso de um dado fenmeno. Desta forma, no importaria se o modelo tem ou no o apoio e a confirmao provenientes de dados empricos, obtidos atravs da manipulao experimental ou da observao naturalista; basta no apresentar falhas lgicas e ter alguma semelhana com o fenmeno que pretende representar.
1.2.
O Mundo e o Laboratrio: A Utilidade de um Modelo
Em um artigo publicado em 1947 (Current Trends in Experimental Psychology ) Skinner elegeu o mtodo experimental como o mais adequado para o estudo cientfico das correlaes funcionais existentes entre o ambiente fsico e social e o comportamento. Segundo Skinner, o comportamento assim como todo e qualquer objeto de estudo de uma cincia natural regido por leis, cujo conhecimento viabiliza a compreenso da imensa variabilidade e multiplicidade nas quais so expressos princpios bsicos de regularidade descritos por estas leis (Lopes Junior, 1993). A vantagem do uso de modelos para tentar explicar fenmenos da Natureza que os mesmos no so contaminados por outros elementos no essenciais compreenso do fenmeno (Morato, 1993). Quanto mais acuradas forem as previses, melhores seriam os modelos, isto , melhor representariam o fenmeno que se pretende compreender (Catania, 1998).
- 74 -
De acordo com Catania (1998), para compreender uma situao precisamos nos livrar dos detalhes que no so essenciais e analis-la. Analisar algo, por sua vez, simplesmente fracion-lo em seus componentes. E, para este propsito, nos voltamos ao laboratrio, comeando com o estudo de organismos mais simples que ns mesmos, em ambientes simples. No caso da Psicobiologia, geralmente um modelo proposto para explicar o comportamento humano, mas a confirmao emprica vem tambm de experimentos com animais. A simplicidade do ambiente de laboratrio pode ainda nos ajudar a ver os variados componentes do fenmeno mais facilmente, pois devemos ser capazes de identificar eventos antes que possamos estudar suas propriedades e o modo mais fcil de faz-lo olhando para o comportamento de organismos no-humanos (Catania, 1998).
1.3.
Os Modelos Experimentais nas Cincias da Sade
Os modelos animais podem ser utilizados em todos os campos da pesquisa biolgica. Na cincia mdica os modelos permitem o melhor conhecimento da fisiologia, da etiopatogenia das doenas, da ao de medicamentos ou dos efeitos de intervenes cirrgicas (Ferreira et al, 2005). A pesquisa pr-clnica, ou seja, com animais, tem sido utilizada principalmente com objetivo de: iniciar a avaliao da segurana, tolerabilidade e possvel eficcia de novas drogas; estudar o mecanismo de ao de psicofrmacos; e auxiliar na elucidao dos mecanismos neurobiolgicos dos transtornos mentais (Andreatini, 2002). Existem diversos modelos experimentais descritos. Em pesquisas na rea da sade os modelos experimentais mais utilizados so as culturas de clulas e tecidos (pesquisa in vitro) e os animais de laboratrio (pesquisa in vivo). Outros modelos experimentais utilizados em pesquisas das reas bsicas da sade tambm podem ser encontrados na Biblioteca Virtual da Sade (Ferreira et al, 2005).
2. PSICOLOGIA EXPERIMENTAL: APRENDIZAGEM
Em todas as espcies, e a todo o momento, se observa diversas experincias influenciando profundamente os mais variados comportamentos. Ns humanos, por exemplo, aprendemos a andar, a falar, a planejar e a amar. Assim como aprendemos a enganar, a odiar e a ter certos medos. Pode-se dizer ento que aprendizado uma base para a melhor adaptao do indivduo, apesar de nem todo aprendizado ser adaptativo. Aprendizagem pode ser definida como um processo por meio do qual um comportamento instalado ou modificado, em funo de um determinado estmulo. Isto , so observadas
- 75 -
mudanas duradouras no comportamento, em resposta s interaes entre o organismo e o seu ambiente. No entanto, para caracterizar-se como aprendizado, essa modificao no deve ser decorrente de tendncias inatas de respostas (instinto ou reflexo), maturao, ou estados temporrios do organismo (como fadiga). Sendo assim, trata-se de um processo que se manifesta por alteraes adaptativas no comportamento individual como resultado da experincia. Tambm importante salientar que o aprendizado no medido diretamente: o que medimos o desempenho. Existem duas formas distintas de aprendizado: 1) Aprendizagem no associativa , na qual o organismo aprende sobre as propriedades de um tipo especfico de estmulo; 2) Aprendizagem associativa, na qual aprendida uma relao entre dois estmulos ou entre um estmulo e uma resposta. Antes de partir para definies dos diferentes tipos de aprendizagem, fundamental definir dois termos que sero amplamente utilizados ao longo deste captulo: ESTMULO e RESPOSTA. Estmulo refere-se a uma parte do ambiente no qual o organismo est inserido e que capaz de afet-lo. Os estmulos podem ser pblicos ou privados: os pblicos so externos ao organismo e os privados ocorrem dentro do organismo, no sendo acessveis a observadores. Os estmulos tm a capacidade de eliciar ou evocar respostas, sendo que estas se referem a componentes do comportamento do sujeito. O comportamento pode ser entendido como a interao entre o ambiente e o organismo, ou, mais especificamente, entre estmulos e respostas.
2.1. Aprendizagem No-Associativa
Habituao: um comportamento adaptativo, observado em todas as espcies - de protozorios a humanos - e que no implica na aquisio de respostas novas e sim na perda de velhas. A habituao ocorre quando um estmulo especfico, que no possui carter de ameaa ao indivduo, apresentado repetidamente, culminando na diminuio ou at mesmo na supresso da resposta inicialmente eliciada por esse determinado estmulo. O comportamento adquirido rapidamente, mediante intervalos pequenos de exposio ao estmulo. No entanto, qualquer alterao no estmulo desencadear as respostas inicialmente observadas, e o sujeito voltar a se comportar em funo do estmulo at que haja nova habituao. Um exemplo quando estamos estudando em um local que possui um ventilador ruidoso. No incio, teremos maior dificuldade para nos concentrarmos e nossa ateno ser voltada ao barulho. No entanto, aps algum tempo expostos a tal rudo, nos habituaremos a ele e poderemos estudar tranquilamente, sem ao menos notar sua presena. Porm, caso o ventilador tenha a velocidade de rotao aumentada de maneira perceptvel (alterao nas propriedades do estmulo), possvel que voltemos a nos incomodar com o rudo, assim como, caso o ventilador seja desligado, notaremos que houve uma mudana no ambiente (notaremos a ausncia do rudo).
- 76 -
Porm, vlido lembrar que nem todos os estmulos apresentados repetidamente levam habituao. muito difcil que esse fenmeno seja observado na presena de um estmulo muito intenso como, por exemplo, o som de um bate-estaca enquanto se estuda. Sob algumas circunstncias, a repetio de estmulo pode levar a uma consequncia oposta da habituao, como a potenciao. Assim, no processo de potenciao, h um aumento na resposta a determinado estmulo depois de repetidas apresentaes deste, sendo mais provvel de ser observada quando o estmulo apresentado tem valor aversivo ou oferece algum risco ao organismo, do que quando este possui carter neutro ou reforador. Por exemplo, choques eltricos geralmente eliciam guinchos em ratos. Se repetidos choques forem apresentados, possvel que ocorra aumento na magnitude da resposta do animal. Outro processo semelhante a sensibilizao. Na sensibilizao, observa-se o aumento do poder eliciador de certos estmulos quando estes so apresentados aps um dado estmulo. Por exemplo, determinados sons eliciam resposta de sobressaltos em ratos, e choques usualmente eliciam guinchos nos animais. Caso seja estabelecido um procedimento no qual o animal recebe um choque, e seguidamente apresentado um som (no associado ao choque), provvel que o sobressalto eliciado pelo som tenha uma magnitude maior do que quando o som apresentado sem a presena prvia do choque. Assim, diz-se que o choque sensibiliza o animal para o som. Desse modo, apesar de no haver nenhuma associao entre a resposta de sobressalto e o choque, esta eliciada em maior magnitude devido sensibilizao causada pelo choque. Outro exemplo quando estamos muito estressados com uma prova, e acabamos por nos irritar com diversas outras situaes. Nesse caso, o estresse da prova nos sensibilizou, e respondemos de modo exacerbado a outros estmulos (no associados prova) em funo dessa sensibilizao.
2.2. Aprendizagem Associativa
Estampagem ou imprinting: aprendizagem baseada em uma nica associao entre estmulos e respostas, com efeitos geralmente irreversveis, em um determinado perodo da vida do organismo, aps o qual no pode mais ser estabelecida. Um exemplo clssico ilustrado por ninhadas de patos que seguem sua me, um tipo de estampagem desenvolvida logo no incio da vida dos filhotes e com carter altamente adaptativo, pois assegura que os filhotes fiquem sempre prximos a ela. Quando um patinho sai da casca do ovo, provvel que a primeira coisa em movimento que ele veja seja sua me, de quem ficar perto e a quem seguir. Todavia, se a me no estiver presente e o patinho vir, de incio, alguma outra coisa em movimento (por exemplo, uma pessoa ou uma mquina), ele ir responder a esse estmulo como teria feito em relao sua me biolgica, seguindo-o.
- 77 -
O etlogo Konrad Lorenz foi um dos pesquisadores que mais se dedicou investigao deste fenmeno, sendo comumente lembrado quando se cita este tipo de aprendizagem. Lorenz costumava estar presente no nascimento de novos patinhos, sendo o primeiro estmulo que os animais entravam em contato, passando a ser seguido por todos os patos. Nesse caso, o estmulo estampado no foi a me pata, e sim o primeiro objeto em movimento visto pelo filhote, o prprio Lorenz. Como dito anteriormente, h um perodo crtico para o desenvolvimento da estampagem. Desse modo, usando o exemplo do patinho, se o estmulo no for apresentado suficientemente cedo na vida do animal, a estampagem pode no ocorrer mais.
2.3. O Comportamento Reflexo ou Respondente (Eliciado)
Todas as espcies animais possuem algumas conexes inatas, chamadas reflexos. Evolutivamente selecionado, o comportamento reflexo fundamental por permitir ao organismo responder adaptativamente a certos estmulos ambientais, sendo estas respostas essenciais para a sobrevivncia do organismo (por exemplo, imagine os danos causados viso se no fosse o mecanismo reflexo de piscar os olhos diante de determinados estmulos). O estudo do comportamento reflexo iniciou-se na fisiologia. No entanto, com a observao de que determinados estmulos e respostas estavam associados em relaes regulares, causais e observveis, os psiclogos tambm passaram a se voltar para o estudo das respostas reflexas, as quais podem ser definidas por meio do seguinte paradigma:
S
Sendo que:
S: o estmulo eliciador, ou ESTMULO INCONDICIONADO (US uncondicioned stimulus), que representa uma mudana ambiental; : pode ser lida como elicia, termo que se refere especificamente s relaes reflexas, ou seja, o mesmo que pelas leis do reflexo, causa, produz; R: chamado resposta reflexa, ou RESPOSTA INCONDICIONADA (UR unconditioned response), estabelecido como a resposta ELICIADA pelo estmulo (S), ou seja, a mudana comportamental causada pelo estmulo.
- 78 -
Esta uma frmula geral e pouco precisa do reflexo, pois no diz sobre como R dependente de S. Assim, dentro do paradigma do comportamento respondente, existem algumas leis que so importantes para compreenso dessa dependncia e regem a regularidade da relao entre os estmulos eliciadores (S ou US) e os respondentes (R ou UR), as quais sero brevemente descritas a seguir. 1) Lei do Limiar: Existe um limite determinado de intensidade do estmulo a partir do qual a resposta sempre ocorrer, e abaixo do qual no ocorrer nenhuma resposta. Assim, preciso que o estmulo tenha certa intensidade para que possa eliciar uma determinada resposta. 2) Lei da Latncia: Entre a apresentao de um estmulo e o incio da resposta, h um pequeno intervalo de tempo, chamado de latncia ou perodo latente. De acordo com a lei da latncia, medida que aumenta a intensidade do estmulo eliciador, diminui o tempo transcorrido (perodo latente) at o aparecimento da resposta eliciada. 3) Lei da Intensidade-Magnitude: medida que aumenta a intensidade do estmulo eliciador, aumenta a magnitude da resposta eliciada. Todas as respostas que podem ser eliciadas em um organismo mantm essa relao de dependncia com a intensidade do estmulo. Os estmulos eliciadores podem ento ser explicitados por meio de um espectro de intensidade , estando a magnitude da resposta relacionada intensidade do estmulo.
3. CONDICIONAMENTO CLSSICO
O processo de condicionamento clssico (ou condicionamento respondente) foi primeiramente estudado no incio do sculo XX pelo fisiologista russo Ivan Petrovich Pavlov, que inicialmente fez observaes bsicas e bastante simples. Pavlov constatou, enquanto realizava estudos sobre o mecanismo fisiolgico da digesto em ces, que certos cidos diludos ou comida posta na boca de ces famintos levava produo de saliva pelas glndulas apropriadas, ou seja, ele observou o reflexo salivar. At a, nenhuma novidade, e de fato no foi essa observao que lhe concedeu seu destaque na psicologia. Alm de observar tal reflexo, Pavlov constatou que saliva tambm era produzia nos ces mesmo antes da comida chegar boca, bastando apenas o animal faminto sentir o cheiro ou ver o alimento. E at mesmo a simples presena do homem responsvel por alimentar o co tambm era capaz de fazer salivar o animal faminto. Estas observaes levantaram questionamentos em Pavlov, que acreditava que certamente essa salivao produzida apenas pela viso da comida ou do alimentador no era um mecanismo inato entre estmulo e resposta, e sim uma relao aprendida de acordo com as experincias vivenciadas pelo animal. Ou seja, havia algum outro elemento alm do estmulo (comida) e resposta (salivao), e esse estmulo o chamado ESTMULO CONDICIONADO (CS, de
- 79 -
conditioned stimulus), estmulo que, precedendo exata e diretamente o estmulo eliciador do reflexo (US), poderia, por si s, eliciar a salivao. Atravs de um experimento clssico, Pavlov conseguiu ento demonstrar a aquisio de novas relaes entre estmulos e respostas, e passou a estudar exaustivamente a generalidade desse paradigma, que veio a ser chamado posteriormente de condicionamento clssico, ou condicionamento respondente, ou mesmo condicionamento pavloviano. No experimento clssico, um co foi colocado em uma sala experimental, e por meio de uma cnula posta na sua mandbula, era possvel quantificar sua resposta de salivar. Por repetidas vezes, era apresentado um som e, logo em seguida, era servido um pouco de alimento ao cachorro. Esse pareamento (ou emparelhamento) foi repetido diversas vezes, por dias seguidos, at que, com o passar do tempo, a prpria apresentao do som era capaz de fazer o animal produzir saliva. Desse modo, no paradigma do condicionamento clssico, h um estmulo previamente neutro (CS ex: um som) que adquire a propriedade de eliciar uma resposta (CR ex: salivar) que originalmente era eliciada por outro estmulo (US ex. comida). E esse processo atravs do qual o CS passa a ter capacidade de eliciar CR aps sucessivos pareamentos com o US o que chamamos CONDICIONAMENTO. Tal paradigma pode ser esquematizado do seguinte modo:
Sendo: CS: um estmulo previamente neutro (som) US: um estmulo incondicionado (comida) UR: uma resposta incondicionada (salivar)
Aps sucessivos pareamentos entre CS-US, tm-se:
No entanto, para que haja essa associao dos estmulos, necessrio um treinamento prvio, por meio do qual ocorra seguidos emparelhamentos entre CS e US, numa relao
- 80 -
temporal de estreita proximidade. Nota-se ento que preciso eliciar uma resposta antes de condicion-la a outro estmulo. importante ressaltar que o condicionamento clssico permite que novos estmulos passem a controlar respostas j existentes, mas no capaz de instalar novas respostas ao repertrio prvio do organismo. O termo resposta condicionada foi assim denominado para que houvesse uma distino da resposta incondicionada, isto , das respostas reflexas ou inatas. Desse modo, resposta condicionada equivale ao termo resposta adquirida, pois se trata de uma resposta adquirida por meio da experincia do organismo (ou seja, por meio do pareamento entre um estmulo incondicionado e um estmulo anteriormente neutro). Seguindo o mesmo raciocnio, falar em resposta incondicionada equivale a falar de respostas inatas, pois so reflexos inatos, que independem de qualquer tipo de treino prvio ou aprendizagem. Assim, no processo do condicionamento clssico, o organismo muda seu comportamento em funo de uma nova contingncia estabelecida entre um estmulo condicionado (CS) e um estmulo incondicionado (US). Esse tipo de associao varia muito em termos de complexidade, e trata-se de um processo muito importante para sobrevivncia do organismo, pois lhe permite adaptao ao meio e s suas frequentes modificaes.
Figura 1: Desenho esquemtico do aparato utilizado por Pavlov em seus experimentos de Condicionamento clssico. Retirado de: www.simplypsychology.pwp.blueyonder.co.uk.
Averso condicionada a sabores - Tipo especfico de condicionamento clssico, tambm conhecido como Efeito Garcia. Trata-se de um processo que envolve reaes fisiolgicas aversivas, como vmito, desconfortos gstricos e mal estar ocasionados aps a ingesto de um alimento. Tal condicionamento tem o diferencial de ser necessria apenas uma exposio ao estmulo (alimento) para que haja o estabelecimento da relao condicionada. Alm disso, o
- 81 -
Efeito Garcia diferencia-se pelo tipo de estmulo envolvido no paradigma, ou seja, envolve apenas condicionamento a estmulos relacionados a alimentos. Por exemplo, dada ao rato uma soluo de gua contendo baunilha (substncia com carter apetitivo ao animal) e veneno no letal. Aps ingerir a gua, o animal apresenta forte reao de vmito. Feito o condicionamento, o animal evitar tomar gua contendo aroma ou sabor de baunilha, mesmo que esteja sem o veneno. Ou mesmo no caso de humanos, suponha que voc coma uma comida que cause fortes dores de estmagos e mal estar. Aps essa ingesto, provvel que apenas o odor ou a visualizao do alimento que provocou tais reaes cause respostas semelhantes s ocorridas anteriormente, quando a comida foi ingerida, como enjo e mal estar.
4. CONDICIONAMENTO OPERANTE
A partir das ideias de Watson, Skinner definiu o comportamento como sendo uma interao entre organismo e ambiente. Ele pretendia provar que qualquer comportamento pode ser aprendido, estudado e explicado seguindo os padres cientficos. Skinner e outros behavioristas da poca mostraram em seus estudos que determinados comportamentos podem ser adquiridos, mantidos e modificados a partir das consequncias que produzem. Resposta e consequncia so termos comumente empregados por eles para designar eventos. Segmentos ou recortes de um determinado comportamento so denominados respostas, e estas normalmente so observveis. J para que a consequncia produza algum efeito sobre a resposta, deve haver uma condio de dependncia entre elas, ou seja, deve ser estabelecida uma relao de contingncia entre resposta e consequncia. Desta forma, a consequncia pode fortalecer ou enfraquecer a probabilidade de que, num momento futuro, a mesma resposta ocorra diante de um estmulo parecido. Quando a consequncia aumenta a probabilidade da ocorrncia da resposta diz-se que uma consequncia reforadora (reforo), mas se por outro lado, esta consequncia diminuir a probabilidade da resposta, diz-se que uma consequncia punitiva (punio). Os processos ditos psicolgicos envolvem interaes entre herana gentica (ou contingncias filogenticas), contingncias ontogenticas (relaes entre o indivduo e as variveis presentes ao longo de sua vida) e as contingncias culturais, ou seja, as relaes decorrentes da vida em sociedade. Este o chamado modelo de Seleo por Consequncias, proposto por Skinner em 1984, a partir da Teoria da Evoluo de Darwin. Por conseguinte, os comportamentos seriam explicados por meio de processos de selees, e no de forma mecnica.
- 82 -
4.1. Paradigma do Condicionamento Operante
Os experimentos com condicionamento operante incluam, portanto, a observao do efeito da consequncia sobre a resposta emitida, ao invs da observao do efeito do pareamento entre dois estmulos. Observe:
Sendo que: S: situao na qual a resposta ocorre R: resposta C: consequncia Em um exemplo tpico de procedimento experimental, o investigador coloca um rato privado de gua em uma caixa de testes (caixas de Skinner) que possui uma alavanca (ou barra) e um bebedouro. Inicialmente, dentro deste ambiente, o animal apresenta respostas (comportamentos) aleatrias (seguindo as tendncias inatas de atividade exploratria). A observao da emisso destas respostas conhecida como nvel operante. As respostas observadas no nvel operante permitem ao investigador comparar os efeitos da liberao da consequncia para uma ou mais dessas respostas, atravs da comparao entre a frequncia das respostas observadas no nvel operante e nas demais fases do experimento. Entretanto, quando o animal no emite por tendncias inatas ou ao acaso a resposta escolhida pelo experimentador, ou seja, quando esta no faz parte do seu repertrio, necessrio realizar o processo de modelagem do comportamento selecionado. Ou seja, aproximaes sucessivas em direo ao comportamento alvo so reforadas at que apenas a resposta selecionada pelo experimentador seja a nica a ser reforada. Assim, em estgios sucessivos, algumas respostas so seguidas por reforo enquanto outras no. Na medida em que a resposta se aproxima da resposta final desejada, os critrios para a liberao do reforo diferencial tambm se modificam, at que a resposta final desejada seja observada. A escolha de um reforador depende da natureza da resposta a ser aprendida e das condies individuais do organismo a ter o comportamento reforado. Assim, a suscetibilidade do organismo ao reforo (probabilidade de ocorrncia) est relacionada a diversas condies, tanto variveis individuais como ambientais. No exemplo do experimento supracitado, geralmente coloca-se o rato em privao de gua ou alimento anteriormente, e o reforo resposta selecionada pelo experimentador (como a presso barra) so gotas de gua ou pelotas de alimento. Seguindo a teoria da evoluo descrita por Darwin no fim do sculo XIX, Skinner ressaltou que a susceptibilidade ao reforo deve-se ao valor de sobrevivncia, enfatizando que a resposta reforada e no o organismo. Por exemplo, a afirmao correta para a resposta de presso a
- 83 -
barra emitida por um rato que tem como consequncia comida : a resposta de presso barra emitida pelo rato foi reforada pela comida e a incorreta: o rato foi reforado pela comida.
Figura 2: Desenho esquemtico da Caixa de Skinner. Retirado de: www.simplypsychology.pwp.blueyonder.co.uk
4.2. Fortalecimento Operante
Na medida em que a resposta passa a ser emitida pelo organismo, esta pode ser fortalecida. Neste processo, ocorre o aumento gradual da frequncia da resposta ao longo do tempo em funo da liberao de uma consequncia reforadora. O efeito que uma consequncia produz sobre o comportamento no fortalecimento operante pode ser verificado pela observao de algumas alteraes que ocorrem na forma do organismo emitir a resposta. Pode-se registrar, portanto, o aumento da taxa de resposta selecionada em relao ao nvel operante; o aumento da taxa desta resposta em relao taxa de outro comportamento que est ocorrendo na situao, as mudanas sequenciais do responder e a variabilidade da topografia da resposta. Topografia refere-se especificidade do movimento realizado na emisso da resposta. Por exemplo, um rato pode pressionar a barra com uma patinha apenas, ou com as duas, ou at mesmo com o rabo. Entretanto importante citar que neste tipo de anlise, normalmente o aspecto relevante o efeito da consequncia sobre o aumento ou diminuio de emisso da resposta, e no necessariamente o tipo de movimento (topografia) envolvido na ao. Ainda em relao ao efeito das consequncias, os reforadores e punidores so classificados em positivos ou negativos.
- 84 -
4.3. Reforo positivo
Os reforadores positivos aumentam a probabilidade de ocorrer a resposta que os antecedem, devido introduo de uma consequncia positiva. Por exemplo: o animal pressiona a barra e obtm um reforo (alimento). Assim, aumenta a probabilidade do animal emitir a resposta de presso barra novamente quando, faminto, for exposto a uma situao semelhante. necessrio ficar claro que no reforo positivo a presena de uma consequncia positiva (agradvel) que mantm a probabilidade de aumento da taxa de respostas.
PARADIGMA DO REFORO POSITIVO
Sendo que: S: situao na qual a resposta ocorre R: resposta S+: estmulo reforador (positivo)
4.4. Reforo Negativo
A remoo de alguns estmulos ambientais exerce efeitos de maneira poderosa no comportamento. O reforo negativo constitui-se na emisso de uma resposta para retirar alguma situao aversiva. Assim, quando uma resposta tem como consequncia esta retirada, tender a aumentar sua probabilidade quando o organismo for exposto a uma situao semelhante no futuro. Por exemplo: o animal pressiona a barra para interromper um estmulo aversivo (choque nas patas). Em um exemplo com humanos, pode ser citado o fato de tomar um medicamento para remover alguma dor.
- 85 -
PARADIGMA DO REFORO NEGATIVO
Sendo que: S-: estmulo aversivo R: resposta So: representa a retirada do estmulo aversivo
4.5. Reforo Negativo e o Comportamento de Fuga e Esquiva
Nestes paradigmas, a resposta de um organismo suspende a presena de um estmulo aversivo. No comportamento de fuga o estmulo aversivo j est presente antes da emisso da resposta que o elimina. No comportamento de esquiva a resposta evita, previne, a ocorrncia do estmulo aversivo. Portanto diferente da fuga j que o estmulo aversivo ainda no est presente no momento em que a resposta de esquiva emitida. Da mesma forma, tanto a resposta de fuga quanto a de esquiva podem ser adquiridas e mantidas no repertrio de um organismo. Como exemplo, temos a situao de correr de uma sala que est pegando fogo. Esta seria considerada uma resposta de fuga. Por outro lado, pode-se correr da sala quando virmos fumaa ou alertas de incndio. Como o fogo (que o estmulo final a ser evitado) ainda no est presente no mesmo local em que o organismo, diz-se que ocorreu uma esquiva.
4.6. Reforo Secundrio ou Condicionado
Estmulos neutros tambm podem adquirir propriedades funcionais de reforadores ou punidores quando so pareados em algum momento da histria do organismo com reforadores primrios ou incondicionados, o que possibilita aos estmulos previamente neutros assumirem a mesma funo dos demais. Os estmulos neutros passam a ser chamados de reforadores secundrios ou condicionados e apresentam a propriedade de sinalizar a ocorrncia do reforo primrio e podem substitu-lo durante o aprendizado. Por exemplo, quando algum ouve uma determinada msica no carro e ocorre um acidente. Se houver um condiciomamento, pode ser que a mesma pessoa passe a apresentar as mesmas respostas fisiolgicas e emocionais de medo (corao acelerado, respirao ofegante e etc) toda vez que ouvir a msica. Assim, um estmulo
- 86 -
que antes era neutro (msica), pareado com um estmulo aversivo (risco sobrevivncia em um acidente de carro), passa a ter a mesma funo que este sobre o organismo.
4.7. Esquemas de Reforamento
Quando toda resposta selecionada seguida de uma consequncia reforadora, podese dizer que o processo de reforamento segue um esquema de reforo contnuo ou simples, isto , uma relao de contingncia do tipo R-S+ prevalece continuamente entre a resposta e o reforo. Porm quando nem toda resposta seguida de uma consequncia reforadora, diz-se que se segue um esquema de reforamento intermitente. Os esquemas de reforamento intermitente produzem efeitos particulares e distintos na resposta do organismo. Para mais detalhes ver Millenson (1967) e Catania (1999). H quatro principais tipos de esquemas de reforamento:
Razo fixa: depende do nmero de resposta que o organismo deve emitir. Este nmero pr estabelecido pelo experimentador. Ex: 10 respostas de presso barra para obter uma consequncia reforadora (alimento). Razo varivel: tambm depende do nmero de respostas apresentadas pelo sujeito, porm o nmero de respostas a serem emitidas para que a consequncia seja liberada varia aleatoriamente. Ex: mquina de caa-nqueis, programada para liberar a recompensa aps um nmero varivel de jogadas. Intervalo fixo: depende do tempo. O reforo s ocorre se a resposta for emitida aps um tempo pr-fixado. Ex: remunerao por hora de trabalho. Intervalo varivel: tambm depende do tempo. Aqui o reforo s se torna disponvel se a resposta ocorrer aps um intervalo de tempo, sendo que este no permanece sempre constante. Ex: tentar falar com uma pessoa ao telefone quando a linha est ocupada.
4.8. Punio
Como foi visto at aqui, o comportamento pode ser adquirido, mantido e modificado por suas consequncias. Porm h uma relao na qual as consequncias da resposta tornam sua ocorrncia menos provvel. Quando uma determinada resposta seguida de uma consequncia punitiva, a probabilidade desta resposta ocorrer novamente diminui. Por exemplo, quando um rato aperta a barra e a consequncia desta resposta um choque nas patas. Nesta operao a
- 87 -
probabilidade da resposta de presso barra diminui na medida em que a consequncia punitiva liberada. A operao de punio consiste em programar para uma determinada resposta uma consequncia aversiva, que tornar a emisso da resposta menos provvel at que seja suprimida. Vale destacar que a resposta punida e no o sujeito, e que a resposta pode reaparecer algum tempo aps a retirada da consequncia aversiva. Seu efeito na resposta depende da intensidade e da repetio da consequncia punitiva, bem como de variveis individuais.
PARADIGMA DA PUNIO
Sendo que: S: estmulo R: resposta S-: estmulo aversivo ou punitivo
4.9. Extino
Diversos comportamentos podem ser adquiridos e mantidos no repertrio do organismo seguindo os paradigmas de reforo descritos at aqui. Porm a probabilidade da resposta pode retornar ao nvel operante ou at desaparecer do repertrio comportamental do organismo pela retirada da consequncia reforadora, ou seja, quando h uma quebra da relao contingencial entre a resposta e a consequncia. Assim, a extino depende de uma nova aprendizagem, visto que a relao entre a resposta e a consequncia deixa de existir. PARADIGMA DA EXTINO
Sendo que: S: estmulo
resposta sem consequncia
- 88 -
Ex: a resposta de bicar um disco, emitida por um pombo, suprimida pela retirada da consequncia (alimento). O comportamento de birra de uma criana no supermercado suprimido pela ausncia de ateno fornecida pela me. Dois dados relevantes referem-se ao fato de que a extino difere do esquecimento porque a resposta pode ser reinstalada quando for novamente reforada, e em certas condies reaparece espontaneamente (recuperao espontnea). O segundo ponto que a taxa de respostas aumenta no incio do procedimento de extino. No raro, pode-se notar um exemplo deste fato na vida real, quando um controle de televiso, por exemplo, deixa de funcionar porque a pilha acabou. Normalmente (mesmo que no notemos), apertamos diversas vezes os botes e at mesmo mudamos a topografia da resposta (passamos a apert-los mais forte, ou de algum modo diferente) at que paramos de responder (apertar os botes).
4.10. Controle de Estmulos: Discriminao e Generalizao de Estmulos
Generalizao definida como o responder de maneira similar a estmulos diferentes, mas que tm alguma propriedade em comum. No incio do processo de condicionamento ocorre resposta generalizada a vrios estmulos fisicamente parecidos como, por exemplo, trs estmulos sonoros (800, 812 e 825 Hz). Por outro lado, discriminao definida como o responder diferencial na presena de estmulos diferentes. Ocorrer discriminao entre os estmulos quando o responder for reforado na presena de somente um deles (no caso do exemplo acima, haver discriminao quando o sujeito passar a responder na presena de apenas um dos trs estmulos sonoros). Ou quando somente um for reforado e o outro for punido.
Condicionamento Clssico Comportamento involuntrio Eliciado pelo estmulo (o comportamento eliciado pelo estmulo, e emitido pelo organismo). Pareamento entre dois estmulos (USCS) Respostas fisiolgicas e emocionais
Condicionamento Operante Comportamento voluntrio Evocado pelo estmulo (o comportamento evocado pelo estmulo e emitido pelo organismo) Associao entre a resposta, um reforo e uma consequncia (S-R-C) Comportamento ativo que leva consequncia
- 89 -
5. MAPAS COGNITIVOS E APRENDIZAGEM LATENTE
A psicologia cognitiva demonstrou que nosso conhecimento do mundo baseado na nossa capacidade biolgica de perceber o mundo, que a percepo um processo construtivo que no depende somente da informao adquirida a partir de um estmulo, mas tambm das estruturas mentais de percepo. Dessa maneira, a psicologia cognitiva no est preocupada simplesmente em como um comportamento particular se estabelece, mas tambm em analisar os processos pelos quais as informaes sensrias so transformadas em percepo e suas consequentes aes. Antes da psicologia cognitiva, acreditava-se que a aprendizagem no ocorria a menos que o comportamento fosse reforado durante o processo de aprendizagem (aprendizagem operante, no esquema S-R-C, estmulo resposta consequncia). Tolman, entretanto, realizou uma srie de experimentos que demonstrou a ocorrncia de outro tipo de aprendizagem. Ele realizava seu trabalho com ratos, utilizando labirintos complexos, como o da figura 3.
Figura 3: Labirinto utilizado por Tolman. Em um dos seus estudos mais importantes, ele investigou qual a importncia que um reforo (ou recompensa) teria na aprendizagem de seus animais no labirinto. Seu experimento usou quatro grupos de animais: Grupo1: Privado de alimento e que recebia recompensa (um
- 90 -
pedao de rao); Grupo2: Privado e sem recompensa; Grupo3: No-privado com recompensa; Grupo4: no-privado e sem recompensa. Tolman testou os animais em seu labirinto e mediu o nmero de erros que os animais apresentavam ao longo dos dias. Segundo as teorias do behaviorismo, a resposta dos animais do Grupo1 em chegar caixa de recompensa mudariam seu comportamento, consequentemente esse comportamento teria maior probabilidade de se repetir. Percebam que os animais do Grupo1 diminuem gradativamente o nmero de erros. Os animais do Grupo1 teriam uma motivao maior, ou drive, para realizar a tarefa mais rpido. Os animais dos outros grupos no teriam uma consequncia ou recompensa favorvel, portanto no teriam porqu de alterarem seu comportamento. E isso o que aconteceu, como podemos ver na figura 4.
Figura 4: Grfico do experimento do lobirinto de Tolman. Ento, para testar, de fato, que a gradual associao estmulo-resposta-consequncia que realiza a aprendizagem, Tolman continou seu experimento. Com um outro grupo de animais, aps dez dias de realizao da tarefa, Tolman resolveu dar recompensa a animais privados que no recebiam recompensa (Grupo2). A partir do dcimo segundo dia, o Grupo 2 comeou a mostrar desempenho semelhante ao Grupo1 (Figura 5). Se o behaviorismo dizia que as alteraes do comportamento so graduais e que exigiam modulao do comportamento atravs de uma recompensa ou consequncia, como os animais do Grupo2 tiveram um aumento de performance to rpido? O experimento sugere que os ratos aprenderam sobre o labirinto durante as corridas iniciais sem recompensa e eles foram capazes de utilizar essas informaes quando a recompensa foi introduzida.
- 91 -
Figura 5: Segunda parte do Experimento de Tolman. O grupo 2, privado e sem alimento, aps receber o alimento demonstra performance to boa quanto a do grupo 1, privado e com recompensa. A partir desses resultados, Tolman formulou a hiptese de Aprendizagem Latente. Ele props que eventos sem significncia aparente podem ser suficientes para gerar aprendizagem. A aprendizagem pode no ser expressa comportamentalmente, mas ela ocorre. O reforo (ou recompensa) afeta o desempenho, medida que elicia a ocorrncia do comportamento decorrente da aprendizagem, mas no necessrio para que a aprendizagem em si ocorra. Quando essa informao necessria, ela resgatada mais facilmente, porque j foi previamente apresentada ao indivduo. Assim, Tolman mostrou que em todo aprendizado ocorre dentro do paradigma da associao S-R-C, e que apenas a apresentao S-S (estmulo-estmulo) j suficiente para que algum aprendizado ocorra. Em seus relatos, Tolman dizia que os ratos privados quando recebiam recompensa apresentavam um desempenho to bom quando os que sempre receberam recompensa, como se eles soubessem o caminho que deveriam fazer e onde a caixa de recompensa estava. Tambm descreve sobre um animal que pulava a parede do labirinto, andava por cima dele e chegava diretamente a caixa onde receberia a recompensa. Aps isso, Tolman desenvolveu o conceito de Mapas Cognitivo. Basicamente, formaramos uma memria espacial quando colocados em um ambiente, mesmo que nesse ambiente no h nada de recompensador. Pense quando voc est em um mercado e lembra no caixa ao final das compras que esqueceu de comprar uma lata de ervilha. Mesmo sem ter uma aprendizagem S-R-C (estmulo lata de ervilha; resposta pegar a lata; consequncia maior probabilidade associar o local com a lata de ervilha), acredito que saber em qual gndola a ervilha est. Pode no saber exatamente, mas saber o local prximo. Outro exemplo quando dirigimos para casa por um trajeto habitual e
- 92 -
encontramos uma das ruas fechadas. Baseando-se na nossa localizao espacial, conseguimos elaborar um caminho alternativo, mesmo sem nunca o termos realizado antes. Em um estudo posterior, Tolman treinou ratos privados de alimento para executar um outro tipo de tarefa. Em um labirinto em T, como da figura abaixo, os animais sempre eram colocados na brao A e recebiam a recompensa no brao B, ou seja, viravam para a direita.
Figura 6: Labirinto em T de Tolman (um dos braos, C ou A, era sempre bloqueado). Esse esquema se repetia por algumas vezes. Em um determinado momento, o labirinto era girado em 180 graus. Agora, os animais eram colocados no brao C. Tolman observou que alguns animais continuavam com o comportamento de virar direita (agora D), ou seja, uma aprendizagem S-R-C virar para a direita e receber uma recompensa. Porm, outros animais viraram para onde estava a recompensa inicialmente, e viravam para a esquerda (mantendo a escolha no ponto B). Esses animais utlizaram a estratgia de mapas cognitvos para gerar uma representao espacial do ambiente virar para o local inicial da recompensa para ganhar a recompensa. Os mapas cognitivos foram os precursores para os conceitos de memria espacial e pensamento espacial. Essas representaes espaciais levam em considerao nossos estados mentais internos, teoria contra as ideias do behaviorismo.
6. MODELOS ANIMAIS PARA ESTUDO DE MEMRIA
Os modelos comportamentais utilizados em estudos de aprendizagem e memria geralmente baseiam-se em teorias da Psicologia Experimental. A maioria dos testes envolvem dois momentos. O momento de aprendizagem, no qual o animal tem contato com as regras da tarefa, e o momento de teste, no qual verifica-se se a aprendizagem da tarefa foi retida pelo animal. Abaixo esto alguns dos modelos comportamentais mais utilizados em estudos de memria.
- 93 -
6.1. Condicionamento de Medo
Essa tarefa um tipo de Condicionamento Clssico Pavloviano que, por conter um contedo emocional forte, aprendida de maneira muito mais rpida e sua memria permanece por muito mais tempo. Nesse modelo, o estmulo incondicionado (US) tem uma caracterstica aversiva e/ou nociva, como um choque nas patas, por exemplo, e elicia respostas reflexas incontrolveis (i.e. pulos e chacoalhar das patas), enquanto o estmulo condicionado (CS) neutro (geralmente um som, Luz ou o prprio ambiente). Aps o pareamento de CS e US, a reapresentao de CS elicia respostas comportamentais e fisiolgicas de medo e ansiadade no animal, como a completa imobilidade do animal (freezing), o alteraes da presso arterial e da frequncia cardaca. Em estudos comportamentais, mede-se o tempo em que o animal permanece imvel e toma-se essa medida de medo como uma medida tambm de memria, partindo-se do princpio que, quanto maior o medo (tempo em congelamento), maior a memria do animal sobre a associao CS-US. Quando o contexto representa o estmulo condicionado, a tarefa depende da integridade do hipocampo e denomina-se condicionamento de medo ao contexto. Por outro lado, quando o estmulo condicionado o som, a tarefa depende do estriado e denomina-se condicionamento de medo ao som. Alm disso, ambas tarefas dependem da integridade da amgdala, uma vez que essa estrutura representa o centro de controle da experincia emocional.
6.2. Esquiva Passiva
Na esquiva passiva (ou inibitria), o animal tem uma resposta especfica (ex, apertar uma barra ou explorar um determinado compartimento da cmara) punida (ex, por um choque). Aps aprender a tarefa, o animal recolocado no ambiente da tarefa e mede-se o tempo (latncia) que o animal leva para realizar o comportamento que foi punido (inibido). Toma-se essa latncia como a medida de reteno da memria da aprendizagem inicial.
6.3. Esquiva Ativa
A esquiva ativa tem dois componetes de aprendizagem. O primeiro um componente uma aprendizagem Pavloviana (associao CS-US), em que se aprende que aps uma determinada sinalizao(CS, ex, som, luz), um evento eversivo ocorrer (US). O segundo uma aprendizagem operante, na qual o animal aprende a exibir uma resposta (esquiva) para evitar o
- 94 -
evento aversivo (geralmente o animal atravessa para uma outra cmara). Aps a aprendizagem dessa tarefa, numa reapresentao do animal cmara e s sinalizaes (CS), mede-se a quantidade de respostas corretas (esquivas bem sucedidas) do animal, e toma-se essas respostas comouma medida de memria.
6.4. Labirinto Aqutico de Morris
O animal colocado em uma piscina com uma plataforma submersa. H uma presso natural do animal a encontrar um local mais seguro e confortvel do que ficar nadando, ento ele nada at encontrar a plataforma. Aps algumas repeties, o animal encontra a plataforma em um tempo menor (at um tempo mnimo), o que considerado uma medida de aprendizagem. No dia do teste, retira-se a plataforma e mede-se o tempo que o animal nada na rea em que a plataforma estava localizada antes. Quanto maior o tempo do animal na regio da plataforma, maior a memria sobre a localizao da mesma. A tarefa se apresenta em duas verses: a espacial e a com pista. Na verso espacial, plataforma est submersa e o animal deve se guiar pelas pistas ambientais para aprender (triangular) sua localizao. Na verso com pista, existe a sinalizao de onde se encontra a plataforma. Assim, o animal s precisa guiar sua resposta baseado em uma nica pista ambiental. Estudos mostram que a leso no hipocampo pode causar prejuzo no desempenho da verso espacial enquanto leses no estriado podem causar prejuzo do desempenho do animal na verso com pista.
6.5. Labirinto em T
Essa tarefa foi desenvolvida por E. C. Tolman, para testar tipos diferentes de aprendizagem. Nessa tarefa, o animal treinado para encontrar um reforo positivo no final de um dos braos de um labirinto em forma de T. Para localizar o brao com a recompensa, o animal pode utilizar como estratgia 1) as pistas espaciais da sala ou 2) a sua prpria movimentao corporal (egocntrica). Em uma segunda parte da tarefa, invert-se a orientao dos braos de modo que o brao que antes era esquerdo, torna-se o direito e vice-versa. Assim, pode-se verificar que tipo de aprendizagem o animal teve, por ex., se a aprendizagem foi espacial, o animal deve virar para o brao correto (que continha a recompensa), se a aprendizagem foi egocntrica, o animal deve fazer o mesmo movimento corporal que fazia antes, o que o leva para o outro brao do labirinto. Estudos sugerem que leses no hipocampo levam a prejuzo no desempenho da estratgia espacial enquanto leses no estriado causam prejuzo na estratgia egocntrica.
- 95 -
6.6. Labirinto Radial
Essa tarefa utiliza um labirinto que possui um centro com oito braos, e se apresenta em duas verses: a win-shift e a win-stay. Na verso win-shift, um reforo positivo colocado ao final de cada brao e os animais devem visitar cada brao apenas uma vez, pois cada entrada em um brao j visitado computada como erro. Na verso win-stay, quatro dos oito braos so iluminados, e apenas esses braos contm a recompensa. Nessa verso, o rato deve aprender a discriminar visualmente o brao do qual ele deve se aproximar. Essa verso envolve a aquisio de memria de hbito, ou aprendizado do tipo estmulo-resposta (S-R). Estudos mostram que leses no hipocampo causam prejuzo no desempenho na verso win-shift enquanto leses no estriado afetam a verso win-stay.
6.7. Reconhecimento de Objetos
Esta tarefa baseada na tendncia de roedores de explorar por mais tempo objetos que so novos, na ocasio de que estes sejam apresentados simultaneamente a objetos familiares. Nessa tarefa, o animal exposto a dois objetos por um determinado tempo. Na sesso de teste, um dos objetos trocado por um objeto novo, diferente dos originais. Nesse caso, se espera que o animal gaste mais tempo explorando o objeto novo do que o objeto familiar. Esse modelo tem a vantagem de utilizar um comportamento natural do animal que a explorao e no envolver um estmulo aversivo.
7. MODELOS ANIMAIS EM TRANTORNOS PSIQUITRICOS
Modelos animais so uma ferramenta essencial para a pesquisa translacional. Essa importncia torna-se ainda mais acentuada em Psiquiatria, uma vez que seu uso permite a maior compreenso das bases neurobiolgicas e dos mecanismos de ao dos agentes teraputicos envolvidas na doena (Andreatini, 2002), alm da avaliao de segurana, tolerabilidade e possvel eficcia de novas drogas (Rodgers et al, 1997). No entanto, a modelagem de transtornos neuropsiquitricos humanos em animais extremamente desafiadora, visto a natureza subjetiva de vrios sintomas, a ausncia de biomarcadores e testes diagnsticos objetivos e a falta de completa elucidao sobre sua etiologia e patologia (Nestler e Hyman, 2010). Assim, modelos animais em doenas mentais
- 96 -
dificilmente
mimetizaro
toda
extenso
de
um
distrbio
neuropsiquitrico
humano,
especialmente como definido atualmente pelo DSM-IVTR (2000). A fim de transpor essa dificuldade e associado variedade de sintomas dos transtornos psiquitricos que podem ser dissecados em suas partes componentes (endofenotipos), muitos autores valem-se da estratgia de usar separadamente modelos animais j existentes para a avaliao de faces distintas da doena (Lenox et al., 2002). Com isto, uma bateria de testes simples, facilmente induzidos e eficientes em avaliar os sinais mais proeminentes da doena pode oferecer um conjunto de ferramentas investigativas mais vlido, abrangente e robusto do transtorno mental. Ainda, com o objetivo de normatizar a fora e legitimidade dos modelos animais, tornou-se til comunidade cientfica padronizar critrios que garantam sua validade de acordo com trs eixos: validade de face, de construo e preditiva (Willner, 1986).
Validade de Face reflete a semelhana anatmica, bioqumica, neuropatolgica e comportamental entre o modelo e a condio patolgica a ser modelada. Validade de Constructo traduz a habilidade do modelo em reproduzir alguns aspectos fisiopatolgicos da doena em estudo. Validade Preditiva revela a eficincias dos agentes teraputicos empregados na clnica em reverter os sintomas induzidos no modelo animal.
Assim, a escolha de um modelo animal adequado para a investigao de determinado fenmeno deve seguir os critrios de validade a fim de aproximar ao mximo as medidas obtidas realidade clnica, bem como diminuir o efeito de variveis que podem dificultar a interpretao e aplicabilidade dos dados. Ressalta-se tambm a importncia do pesquisador pautar o procedimento experimental em princpios ticos a fim de minimizar o sofrimento dos animais.
7.1. Modelos animais de Transtornos de Ansiedade
Apesar da plena expresso de reaes de ansiedade ser um fenmeno tipicamente humano, possvel encontrar correlaes entre os transtornos de ansiedade e respostas de defesa que animais exibem frente a estmulos ou situaes de perigo. Com isto, vrios estudos tm definido a ansiedade como componente essencial de um sistema neural que vem se mantendo ao longo da evoluo das espcies. Esse sistema responsvel no apenas pela deteco de sinais de perigo no meio ambiente, mas tambm pela ativao de reaes fisiolgicas e comportamentais adequadas de defesa. Neste contexto, os modelos animais de ansiedade so utilizados para investigar os diversos padres etolgicos associados doena, bem como delinear os mecanismos neuroqumicos
- 97 -
envolvidos nela. Dois modelos muito utilizados na avaliao dos comportamentos emocionais de medo e ansiedade so fundamentados no comportamento natural dos roedores em evitar situaes potencialmente perigosas labirinto em cruz elevado (LCE) e campo aberto, e ambientes iluminados caixa claro-escuro. Labirinto em cruz elevado (LCE) este teste consiste colocar o animal em um labirinto elevado do solo formado por dois braos fechados por paredes e dois abertos (Figua 1), onde se analisa principalmente a frequncia de entradas e o tempo gasto em cada tipo de brao. Considera-se um ndice fidedigno de ansiedade a porcentagem da preferncia pelos braos abertos ou fechados, em que quanto menos o tempo de permanncia e nmero de entradas nos braos abertos, maiores os nveis de ansiedade (Handley & Mithani, 1984). Caixa claro-escuro Trata-se de uma caixa experimental dividida em dois
compartimentos, sendo um deles iluminado e outro no. Diante da averso natural dos animais a ambientes iluminados, seus nveis de ansiedade podem ser estipulados pela preferncia entre os compartimentos, em que o menor tempo de permanncia e nmero de entradas no ambiente iluminado indicaria maiores nveis de ansiedade (Crawley e Goodwin, 1980; Figura 2).
Figura 7: Labirinto em cruz elevado
Figura 8: Caixa claro-escuro
7.2. Modelos Animais de Depresso
Embora os estudos com neuroimagem tenham proporcionado grandes avanos na compreenso dos circuitos neurais do humor e uma srie de distrbios neuroqumicos e neuroendcrinos tenham sido descritos em indivduos deprimidos, nenhuma anormalidade provou-se suficientemente robusta ou consistente o suficiente para diagnosticar a depresso em seres humanos ou validar um modelo animal (Krishnam e Nestler, 2008). Alm disto, variantes genticas altamente penetrantes que causam depresso ainda no foram identificadas. Estas consideraes sublinham o desafio na construo e validao de modelos animais de depresso. Assim, muitos modelos animais tm sido validados para o estudo do comportamento depressivo a partir da observao de que o estresse e a perda emocional so potentes fatores de
- 98 -
risco esta doena. Em geral, os animais so expostos aguda ou cronicamente a um evento estressante inescapvel como reteno, choque nas patas ou temperaturas baixas. Posteriormente, esse animal submetido ao mesmo contexto do evento estressante e comportamentos passveis de serem associados depresso, como a perda da sensao de prazer (anedonia) evidenciada no teste de preferncia de sacarose, e que podem ser revertidos pelo tratamento com antidepressivos so avaliados. Dentre os modelos para estudo da depresso em roedores, o teste do nado forado e o desamparo aprendido so os mais utilizados por apresentarem validade preditiva relevante. Teste do Nado Forado Proposto por Porsolt e colaboradores (1977), baseia-se na exposio de roedores a uma situao aversiva referente a nadar em um tanque cilndrico com gua, onde eles no podem tocar seu fundo ou fugir (escape). Com o tempo, os animais percebem que no tem como sair do local e desenvolvem uma postura de imobilidade. O teste filmado e avaliam-se os perodos de imobilidade e natao, parmetros responsivos ao tratamento com antidepressivos e sensveis validao preditiva de drogas com este potencial (Figura 3).
Figura 9: Nado Forado.
Desamparo Aprendido neste modelo proposto por Seligman (1975), roedores so expostos a situaes de eventos incontrolveis, no contingentes (estmulos aversivos) como choques eltricos nas patas, gerando reaes de desamparo (Figura 4). Isto impede que estes animais aprendam uma resposta de fuga e esquiva (aprendizagem operante). Assim, maiores latncias das respostas de fuga e esquiva ou a no aprendizagem dessas respostas, caracterizam o desamparo.
- 99 -
Figura 10: Desamparo Aprendido.
Outros modelos apoiados no estresse psicossocial, como os testes de suspenso da cauda, derrota social (confronto), isolamento social (separao materna) tambm so empregados. Ainda, vrios procedimentos tm alterado a homeostase de glicocorticides do animal, baseados nos distrbios do eixo HPA envolvidos na depresso, seja pelo seu tratamento crnico ou pela expresso anormal de recepetores deste hormnio em roedores mutantes como forma de modelar a depresso.
7.3. Madelos Animais de Transtorno Bipolar
desenvolvimento
de
modelos
animais
adequados
ao
Transtorno
Bipolar
particularmente desafiador, j que alm de todos os obstculos comuns em modelar os transtornos psiquitricos, ainda existe o carter oscilatrio da doena. Neste sentido, pesquisadores tm se esforado para desenvolver modelos que eliciem tanto caractersticas da fase relacionada depresso, quanto da mania. Na prtica, diferentes testes so utilizados para abordar estes polos da doena, sendo a fase depressiva estudada com os modelos animais supracitados. A observao de que psicoestimulantes podem produzir sintomas semelhantes mania em indivduos saudveis, bem como exacerbar ou induzir episdios manacos naqueles acometidos pela doena (Meyendorff et al, 1985) levou a utilizao da induo farmacolgica como uma ferramenta til para modelar a mania. Por ser um parmetro objetivo, facilmente induzvel e mensurvel, a hiperatividade o aspecto da doena enfatizado sendo avaliada por caixas automatizadas de atividade motora ou por campo aberto (Figura 5).
- 100 -
Figura 11: Campo Aberto.
Assim, modelos animais de hiperatividade como comportamento relacionado mania induzidos por anfetamina ou pelo inibidor da bomba Na +/K+ (ouabana) esto bem estabelecidos, uma vez que apresentam validade de face - o aumento de atividade exibido por pacientes acometidos pelo transtorno bipolar, e validade preditiva - estabilizadores de humor como o ltio e o valproato diminuem a hiperatividade induzida (el-Mallakh et al., 1995; Frey et al., 2006; Kato et al, 2007; Riegel et al, 2009; Jornada et al, 2010). Atualmente, manipulaes genticas como a superexpresso de glicognio sintase quinase-3 (GSK3) e a subexpresso no gene Clock tambm so empregadas como modelo animal de mania por suas correlaes neurobiolgicas com a doena.
7.4. Modelos Animais de Esquizofrenia
A complexidade do quadro de esquizofrenia, seja em termos de manifestao clnica seja por sua etiologia, dificulta o desenvolvimento de modelos animais para este distrbio. Portanto, a validade de muitos dos modelos existentes repousa, quase unicamente, no critrio de validade preditiva. o caso do teste de catalepsia, de estereotipia e a esquiva inibitria, em que se identifica a capacidade do neurolptico ou outros agentes com potenciais antipsicticos de antagonizar receptores dopaminrgicos, revertendo ou atenuando assim os comportamentos eliciados. Objetivando explorar caractersticas circunscritas da doena, como o dfict de processamento de informaes, os modelos atualmente mais empregados so a inibio prpulso do reflexo do sobressalto, teste de atividade locomotora e exploratria, interao social e inibio latente, alm de modelos de leso de determinadas estruturas para modelar a psicose em animais, alteraes neuroqumicas pela administrao de drogas psicomimticas e genticos com camundongos transgnicos.
- 101 -
Inibio pr-pulso (teste do sobressalto) o teste realizado em caixa de isolamento acstico (Figura 6) onde o pr-pulso - um estmulo fraco geralmente acstico, apresentado antes do pulso - um estmulo forte tambm acstico. A inibio por pr-pulso do reflexo do sobressalto est reduzida em animais de laboratrio sob efeito de drogas psicotomimticas, quando o pr-pulso perde a capacidade de inibir a resposta do sobressalto causada pelo pulso. Isto indica incapacidade do animal prestar ateno ao pr-pulso, pelo comprometimento do circuito crtico-estriado-plido-pontino, responsvel pela filtragem da informao sensorial ou cognitiva.
Figura 12: Teste de Sobressalto. Inibio latente teste do efeito deletrio sobre a aquisio de condicionamento clssico provocado pela pr-exposio a um estmulo anteriormente utilizado como estmulo condicionado (aprendizagem associativa). Este fenmeno apresenta-se atenuado ou ausente em pacientes esquizofrnicos na fase aguda devido aos prejuzos cognitivos inerentes doena. Sendo assim, drogas agonistas dopaminrgicas (ex: anfetamina) reduzem a inibio latente e bloqueadores de receptores dopaminrgicos (ex: clorpromazina) a fortalecem.
8. MODELOS EXPERIMENTAIS PARA ESTUDOS DE DEPENDNCIA
8.1. Modelo de Auto-Administrao
Estes modelos apresentam uma alta validade para fins de comparao com consumo humano, por isso so muito utilizados em pesquisas pr-clnicas. A caracterstica fundamental desse modelo de que aps um procedimento de treino o animal passa a auto-administrar a droga de interesse. De uma perspectiva comportamental, os procedimentos de autoadministrao podem ser do tipo operante ou no operante. O modelo operante segue uma aprendizagem operante do tipo S-R-C. Na maior parte dos procedimentos, o animal deve apertar uma alavanca ou enfiar seu focinho em um local especfico para que receba a droga. A via de
- 102 -
administrao muito importante nesse contexto, pois define todo o protocolo experimental. As principais variveis observadas em modelos operantes so ligadas ao comportamento em si como frequncia e taxa de ocorrncia para tal so utilizados modelos de razo fixa e razo progressiva.
Figura 13: Esquema representando a auto-administrao operante em um modelo de roedor. Retirado de Sanchis-Segura & Spanagel, 2006
Em roedores, o modelo no operante utilizado o da auto-administrao oral, sendo majoritariamente utilizado na pesquisa com etanol, pois outras drogas de abuso como cocana, morfina e nicotina. No entanto, estas outras drogas apresentam um baixo potencial motivador quando administradas dessa maneira. Uma caracterstica muito relevante desse modelo, e que aumenta sua validade, o fato do indivduo poder escolher entre consumir etanol ou no, no momento que quiserem, similar ao que observado em humanos. Nesse procedimento, duas garrafas so disponibilizadas ao animal, uma contento etanol na concentrao definida pelo avaliador e outra com gua. As variveis observadas nesses procedimentos esto relacionadas com a quantidade de etanol consumida (expressas em gramas de etanol / kg de peso corporal / tempo), preferncia etanol sobre gua e quantidade de fluido ingerida. Procedimentos em que o
- 103 -
tempo que o animal tem para consumo limitado, como Drinking in the Dark (DiD), j demonstraram ser mais eficazes e menos sujeitos a vieses.
8.2. Preferncia Condicionada Ao Lugar
Os modelos de preferncia condicionada seguem o paradigma Pavloviano de condicionamento clssico. Os efeitos da droga utilizada (US) sero repetidamente pareados com um estmulo neutro do ambiente que aps as sesses de condicionamento se tornar o estmulo condicionado (CS). Nessa questo, o modelo de dependncia mais utilizado a preferncia condicionada ao lugar (do ingls, conditioned place preference CPP). Neste procedimento, o animal exposto a um aparato que consiste de dois ou trs compartimentos. Dois compartimentos apresentam pistas sensoriais que os diferem, sendo estes os estmulos neutros iniciais. Inicialmente, a preferncia inicial do animal deve ser avaliada, para tal o animal tem livre acesso por todo aparato, e o tempo gasto em cada compartimento mensurado, esta etapa chamada de habituao. Este passo crucial para a definio do protocolo experimental a ser seguido. Aps essa etapa, tem incio o pareamento, na qual, em sesses alternadas, os efeitos da droga so pareados em um compartimento e no oposto uma droga incua pareada. Nesta etapa, o animal confinado no compartimento escolhido. Seguida dessa etapa, ocorre o teste de preferncia condicionada, sendo este semelhante habituao, no entanto, espera-se que o animal passe mais tempo no compartimento no qual foi pareado com os efeitos da droga.
- 104 -
Figura
14:
Esquema
ilustrativo
do
condicionamento de preferncia de local em um modelo de roedor. Retirado de Sanchis-Segura &
Spanagel, 2006.
8.3. Auto-Estimulao Eltrica Intracraniana
Este modelo foi essencial para o estabelecimento de uma base neural para a funo de recompensa, pois demonstra que a ativao direta de reas cerebrais especficas de animais capaz de iniciar e manter comportamentos de auto-estimulao (eltrica). Basicamente, neste protocolo o animal tem um eletrodo implantado em reas associadas recompensa. Por meio desse eletrodo, estimulao eltrica (corrente) aplicada para reforar e sustentar um comportamento (ex: pressionar uma alavanca), de maneira que o animal voluntariamente repete o comportamento para receber a estimulao eltrica, que a prpria recompensa. Cada animal pode ter um limiar prprio para a auto-estimulao (ou limiar de recompensa), que corresponde corrente ou freqncia de estimulao mnimas para que o animal mantenha o comportamento de pressionar a alavanca, por exemplo. Uma caracterstica abordada nessa questo de que se administradas anteriormente, diferentes drogas de abuso conseguem reduzir esse limiar, demonstrando uma clara associao entre os efeitos da droga e a estimulao eltrica na regio de interesse, e sugerindo que os efeitos de recompensa das drogas tambm seriam mediados por este mesmo circuito neural.
- 105 -
Figura 15: Esquema ilustrativo da auto-estimulao eltrica intracraniana em modelo de roedor. Retirado de Sanchis-Segura & Spanagel, 2006.
8.4. Sensibilizao Comportamental
A sensibilizao comportamental refere-se ao aumento do efeito estimulante sobre a atividade locomotora produzida por drogas de abuso aps administraes repetidas. Em modelos experimentais, este efeito pode ser observado atravs do aumento progressivo da atividade locomotora, estereotipia ou da atividade rotacional, aps administraes de diferentes drogas de abuso. Uma caracterstica neurobiolgica que embasa esse modelo de que a circuitaria envolvida neste fenmeno homloga quela subjacente aos efeitos de reforo. As neuroadaptaes subjacentes sensibilizao comportamental s drogas de abuso alterariam o funcionamento do circuito de recompensa cerebral sendo, possivelmente, um fenmeno crtico na transio entre o uso moderado e abusivo dessas substncias. O procedimento adotado neste modelo envolve duas fases: induo ou desenvolvimento, que ocorre durante o tratamento com a droga. Esta fase seria o momento em que ocorrem as alteraes celulares e moleculares iniciais, que poderiam alterar de forma permanente a funo neural. A outra fase a expresso (curto ou
- 106 -
longo prazo) na qual so observadas as alteraes neurais permanentes e duradouras que so decorrentes da administrao repetida da droga. Para verificar a expresso da sensibilizao comportamental, desafios com a droga so realizados em todos aps um perodo de abstinncia, posterior ao tratamento. Em geral, manipulaes farmacolgicas (administrao de agonistas ou antagonistas de receptores envolvidos) ou leses na rea Tegmental Ventral (VTA) alteram o desenvolvimento ou induo, e quando feitas no ncleo accumbens afetam a fase de expresso da sensibilizao comportamental.
Figura
16:
Esquema
ilustrativo
da
sensibilizao comportamental em um modelo de roedor. Retirado de Sanchis-Segura &
Spanagel, 2006
- 107 -
REFERNCIAS UTILIZADAS E LEITURAS SUGERIDAS
Andreatini R. (2002) A importncia dos modelos animais em psiquiatria. Rev. Bras. Psiquiatr. So Paulo: Oct, , vol.24, n.4. Braisby, N.; & Gellatly, A. (2005) Cognitive Psychology. New York: Oxford University Press. Cameschi, C. E., & Abreu-Rodrigues, J. (2005). Contingncias aversivas e comportamento emocional. In J. Abreu-Rodrigues & M. R. Ribeiro (Eds.), Anlise do Comportamento: Pesquisa, Teoria e Aplicao (pp. 113-137). Porto Alegre, RS: Artmed. Catania, A. C. (1999). Aprendizagem: comportamento, linguagem e cognio. 4 ed. Porto Alegre: Artes Mdicas Crawley, J.N., Goodwin, F.K. Preliminary report of a simple animal behaviour for the anxiolytic effects of benzodiazepines. Pharmacol Biochem Behav. 1980; 13: 167 170. Deese, J.; Hulse, S. H. The Psychology of learning. 3 ed. Mac Graw-Hill Book Company, 1967. DSM-IV-TR Diagnostic and Statistical Manual of Mental Disorders. 2000; Fourth edition, Text revision. USA, American Psychiatric Association. el-Mallakh RS, Harrison LT, Li R, Changaris DG, Levy RS. An animal model for mania: preliminary results. Prog Neuropsychopharmacol Biol Psychiatry. 1995; 19 :955962. Ferreira, L. M.; Ferreira, L.R. (2003) Experimental model: Historic and conceptual revision. Acta Cir. Bras. So Paulo: , v.18 n.spe. Ferreira, L.M. et al. (2005) Modelos experimentais em pesquisa. Acta Cir. Bras. So Paulo: vol.20, suppl. 2. Frey BN, Valvassori SS, Reus GZ, Martins MR, Petronilho FC, Bardini K, et al. Effects of lithium and valproate on amphetamine-induced oxidative stress generation in an animal model of mania. J Psychiatry Neurosci. 2006; 31: 326332. Garrett, H.E. Grandes experimentos da Psicologia. So Paulo. Cic. Editora Nacional, 1959. Guimares, R. P. (2003). Deixando o preconceito de lado e entendendo o Behaviorismo Radical. Psicologia: cincia e profisso, 23 (3), 60-67. Handley SL, Mitani S. Effects of alfa-adrenoceptor agonists and antagonists in a maze exploration model of fear-motivated behavior. Naunyn-Schmiedebergs Arch Pharmacol. 1984; 327: 15. Harrym F Harlow; James L.McGaugh; Richard F. Thompson (1971) Psychology Albion Publishing company: So Francisco, Califrnia. Jornada L. K., Moretti M., Valvassori S. S., Ferreira C. L., et al. Effects of mood stabilizers on hippocampus and amygdala BDNF levels in an animal model of mania induced by ouabain. J. Psychiatr. Res. 2010; 44: 506510. Kato T, Kubota M, Kasahara T. Animal models of bipolar disorder. Neurosci Biobehav Rev. 2007; 31: 832842. Lazerson, A. Conditioning and learning. In:____ Psychology Today: an introduction. 2 ed. Del Mar, California. CRM Books, 1972 p. 39-59. Lenox RH, Gould TD e Manji HK. Endophenotypes in bipolar disorder. Am J Med Genet.
- 108 -
2002;114: 391-406. Lopes Jr, J. (1993) Consideraes conceituais acerca do behaviorismo radical e da anlise (experimental) do comportamento. In: Nascimento Jr, A.F. (org..) Anais de Etologia, 11. Bauru: UNESP,. p. 13-20. Kantowitz, B. H.; Roedinger III, H. L.; & Elmes, D. G. (2006) Experimental Psychology. Monterey: Thompsom/Wadsworth. Machado, L. M. C. M. (1986). Esquemas de reforamento positivo: esquemas simples. Psicologia, 12 (2), 1-15. Mackintosh, N.J. Discrimination Learning. In: ____ The Psychology of Animal Learning. London. Oxford Univ. Press, 1974 p. 579-583. Mackintosh, N.J. Laws of Association. In: ____ Conditioning and Associative Learning. New York. Oxford Univ. Press, 1983 p. 222-239. Manning, A. Introduo ao comportamento animal. Rio de Janeiro. Livros Tcnicos e Cientficos Editora S/C, 1977. Meyendorff E, Lerer B, Moore NC, Bow J, Gershon S. Methylphenidate infusion in euthymic bipolars: effect of carbamazepine pretreatment. Psychiatry Res. 1985; 16 :303308. Millenson, J.R. (1976). Princpios de anlise do comportamento. Braslia: Coordenada Morato, S. (1993) Modelos Psicobiolgicos aplicados a humanos. In: Nascimento Jr, A.F. (org.). Anais de Etologia, 11. Bauru: UNESP,. p. 21-31. Nestler, E. J., and Hyman, S. E. (2010). Animal models of neuropsychiatric disorders.Nat. Neurosci. 2010; 13: 11611169. Nestor A. Schmajuk, (1997) Animal learning and cognition A neural network approach.Cambridge University Press. , 1997, NY- USA. O'Keefe, J. & Nadel, L. (1978) The hippocampus as a cognitive map. Oxford. Oxford University Press. Olton, D.S. (1979) Mazes, maps and memory. American Psychologist, 34(7): 583-596. Popper, K. (1993) A Lgica da Pesquisa Cientfica. So Paulo: Cultrix. Porsolt R D, Le Pichon M & Jalfre M. Depression: a new animal model sensitive to antidepressant treatments. Nature. 1977; 266 :730-2. Riegel RE, Valvassori SS, Elias G et al. Animal model of mania induced by ouabain: evidence of oxidative stress in submitochondrial particles of the rat brain. Neurochem Int. 2009; 55:491495. Rodgers RJ, Cao B-J, Dalvi A, Holmes A. Animal models of anxiety: an ethological perspective. Brazilian J Med Biol Res 1997; 30: 289-304. Sanchis-Segura, C. & Spanagel, R. (2006). Behavioural assessment of drug reinforcement and addictive features in rodents: an overview. Addiction Biology, 11, 2 38. Schultz, D. P. & Schultz, S. E. (2004). Histria da Psicologia Moderna.Thomson Learning (Pioneira), So Paulo. Seligman, Martin E. P. Helplessness: On Depression, Development, and Death. San Francisco: W.H. Freeman. 1975. Skinner, B. F. (1974). Sobre o Behaviorismo. Cultrix. So Paulo
- 109 -
Skinner, B.F. (2003). Cincia e Comportamento Humano. Martins Fontes, So Paulo. Souza Jr., E. J. & Cirino, S. D. (2004). Esquemas de reforamento. In C. E. Costa; J. C. Luzia & H. H. N. Sant'Anna (Org.). Primeiros Passos em Anlise do Comportamento e Cognio. Santo Andr: ESETec Souza, D. G. (2000). O conceito de contingncia: um enfoque histrico. Temas em Psicologia da SBP, 8(2), 125-136. Squire, L.R.; Kandel, E.R. Brain systems from declarative memory. In:____ Memory: From Mind to Molecules. New York. Scientific American Library, 1998 p. 83-107. Todorov, J. C. (1985). O conceito de contingncia trplice na anlise do comportamento humano. Psicologia: Teoria e Pesquisa, 1(1), 75-87. Todorov, J. C. (1989). A psicologia como o estudo de interaes. Psicologia: Teoria e Pesquisa, 5(3), 347-356. Tolman, E.C. - Cognitive maps in rats and men. Psychol. Ver., 55: 189-208, 1948. http://www.yorku.ca/dept/psych/classics/tolman/maps/maps.htm Tolman, E.C.; Ritchie, B.F.; KALISH, D. - Studies in spatial learning: II. Place learning versus exponse learning. J. Exp. Psychol., 35: 221-229, 1946. Watson, J. B. (1913). Psychology as the Behaviorist Views it. Psychological Review, 20, 158-177. Willner, P. (1986) Validation criteria for animal models of human mental disorders: learned helplessness as a paradigm case. Prog. Neuro-Psychopharmacol. & Biol. Psychiat. Gr-Bretanha:, v.10, p. 677-690.
- 110 -
Vous aimerez peut-être aussi
- 7 Chakras do corpo humanoDocument46 pages7 Chakras do corpo humanoRosi Lopes80% (5)
- Cânone A Duas Vozes PDFDocument8 pagesCânone A Duas Vozes PDFArnaldo Oliveira AlvesPas encore d'évaluation
- Instalação e Configuração de Sistemas OperativosDocument211 pagesInstalação e Configuração de Sistemas OperativosAna Cristina Colaço DiasPas encore d'évaluation
- Extração de Dna de MorangoDocument10 pagesExtração de Dna de MorangoJean Bressan AlbarelloPas encore d'évaluation
- Apostila 3 - 2013Document134 pagesApostila 3 - 2013Marcus Vinicius AlvesPas encore d'évaluation
- Exercicio - Escala de Coma de GlasgowDocument2 pagesExercicio - Escala de Coma de GlasgowJosé Silva0% (1)
- Apostila 2 - 2013Document113 pagesApostila 2 - 2013Marcus Vinicius Alves100% (1)
- Alves 2016 SBNP Boletim - JunhoDocument4 pagesAlves 2016 SBNP Boletim - JunhoMarcus Vinicius AlvesPas encore d'évaluation
- Rivero Et Al 2016 - Player Exp Questionnaire - I J Psych NeuroDocument29 pagesRivero Et Al 2016 - Player Exp Questionnaire - I J Psych NeuroMarcus Vinicius AlvesPas encore d'évaluation
- Alves 2012 SBNP Boletim - MaioDocument2 pagesAlves 2012 SBNP Boletim - MaioMarcus Vinicius AlvesPas encore d'évaluation
- Qualidade de Vida em Idosas: A Importância Da Dimensão SubjetivaDocument8 pagesQualidade de Vida em Idosas: A Importância Da Dimensão SubjetivaMarcus Vinicius AlvesPas encore d'évaluation
- Apostila CV 2009Document158 pagesApostila CV 2009Marcus Vinicius AlvesPas encore d'évaluation
- Pereira, Estramiana, Alves - 2010 - Crenças Essencialstas Sobre Policiais e Delinquentes - Psicol Teor PesqDocument9 pagesPereira, Estramiana, Alves - 2010 - Crenças Essencialstas Sobre Policiais e Delinquentes - Psicol Teor PesqMarcus Vinicius AlvesPas encore d'évaluation
- Estereotipos Automatismos Controle e A Identificacao de Armas e Ferramentas em Diferentes ContextosDocument23 pagesEstereotipos Automatismos Controle e A Identificacao de Armas e Ferramentas em Diferentes ContextosMarcus Vinicius AlvesPas encore d'évaluation
- Leonardo Marona - Pequenas Biografias Não AutorizadasDocument90 pagesLeonardo Marona - Pequenas Biografias Não Autorizadasjubscaldas10Pas encore d'évaluation
- Consolidação Das Leis Do Trabalho (CLT) - DL 5.453.43Document193 pagesConsolidação Das Leis Do Trabalho (CLT) - DL 5.453.43Tâmara Monteiro100% (1)
- O currículo entre o conhecimento e a opiniãoDocument22 pagesO currículo entre o conhecimento e a opiniãoFernanda Binda Alves TouretPas encore d'évaluation
- A CABALA, A MAÇONARIA E OS CARGOS E LOJA - Dermivaldo CollinettiDocument2 pagesA CABALA, A MAÇONARIA E OS CARGOS E LOJA - Dermivaldo CollinettiJoaoCRLMPas encore d'évaluation
- Ideia - História Da ArteDocument68 pagesIdeia - História Da ArtelismorilaPas encore d'évaluation
- Recibo de pagamento SescDocument1 pageRecibo de pagamento SescjfmantovaPas encore d'évaluation
- DIREITO ADM BRASILEIRO: PRINCÍPIOS E NOÇÕES INICIAISDocument77 pagesDIREITO ADM BRASILEIRO: PRINCÍPIOS E NOÇÕES INICIAISshinewellPas encore d'évaluation
- Coerência e coesão no textoDocument3 pagesCoerência e coesão no textotao do somPas encore d'évaluation
- Historia Mocambinho Teresina - 1397523753 ARQUIVO DjalmaFilho ReparadoDocument20 pagesHistoria Mocambinho Teresina - 1397523753 ARQUIVO DjalmaFilho ReparadoEjohansson Veras100% (1)
- Transexualidade Saude Publica e AtuacaoDocument30 pagesTransexualidade Saude Publica e AtuacaoPaulo Sérgio RavanelliPas encore d'évaluation
- Ografia 5ano Manha 1bimDocument6 pagesOgrafia 5ano Manha 1bimDalva Cecília Soares100% (1)
- Estabilidade e Controle - FACULDADE DE CIÊNCIAS AERONÁUTICASDocument32 pagesEstabilidade e Controle - FACULDADE DE CIÊNCIAS AERONÁUTICASChefiaPas encore d'évaluation
- EFEITOS DAS FOLHAS, CASCAS, RAÍZES E FRUTOS DA AMOREIRA (Morus Nigra L.) UTILIZADOS COMO FITOTERÁPICO NA MEDICINA POPULAR PDFDocument4 pagesEFEITOS DAS FOLHAS, CASCAS, RAÍZES E FRUTOS DA AMOREIRA (Morus Nigra L.) UTILIZADOS COMO FITOTERÁPICO NA MEDICINA POPULAR PDFAna LidiaPas encore d'évaluation
- Comparativo de CRMs - Preços, Integrações e ComunicaçõesDocument1 pageComparativo de CRMs - Preços, Integrações e ComunicaçõesBIM RegularizaçõesPas encore d'évaluation
- Mensagem Devocional - Romanos 13Document1 pageMensagem Devocional - Romanos 13JuristanaredePas encore d'évaluation
- Apontamento de Uma Plano de NegociosDocument4 pagesApontamento de Uma Plano de NegociosluciaPas encore d'évaluation
- Ciências avaliação diagnósticaDocument4 pagesCiências avaliação diagnósticaWELDER WILIAN DA SILVAPas encore d'évaluation
- Diário de um Educador: Refletindo sobre o papel do educadorDocument14 pagesDiário de um Educador: Refletindo sobre o papel do educadorjoaquimPas encore d'évaluation
- Conte Comigo HINODocument3 pagesConte Comigo HINOMaria SilvaPas encore d'évaluation
- Timothy Rice - Ethnomusicology in Times of Trouble (2014)Document20 pagesTimothy Rice - Ethnomusicology in Times of Trouble (2014)Mauro MouraPas encore d'évaluation
- Avaliação de riscos profissionais na limpeza de vidro usando o método de William FineDocument44 pagesAvaliação de riscos profissionais na limpeza de vidro usando o método de William FineCarlitos Branquinho KbyPas encore d'évaluation
- Resumo Das Teorias Do DesenvolvimentoDocument1 pageResumo Das Teorias Do DesenvolvimentoAnderson NascimentoPas encore d'évaluation
- Pudim de Leite Condensado MOÇA - Receitas NestléDocument3 pagesPudim de Leite Condensado MOÇA - Receitas Nestlékenia cristinaPas encore d'évaluation
- Analise Da Cations e AnionsDocument6 pagesAnalise Da Cations e AnionsKarolWalcanaiaPas encore d'évaluation
- Apostila Do Curso Introducao Ao Estudo Da Anatomia HumanaDocument35 pagesApostila Do Curso Introducao Ao Estudo Da Anatomia Humanamateusandrea3Pas encore d'évaluation