Vous aimerez peut-être aussi
- Type of ExecutiveDocument1 pageType of ExecutiveArun MauryaPas encore d'évaluation
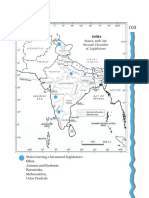
- Chapter 5: Legislature: States With The Second Chamber of LegislatureDocument1 pageChapter 5: Legislature: States With The Second Chamber of LegislatureArun MauryaPas encore d'évaluation
- FPTP & PR SystemDocument1 pageFPTP & PR SystemArun MauryaPas encore d'évaluation
- Directive PrincipleDocument1 pageDirective PrincipleArun MauryaPas encore d'évaluation
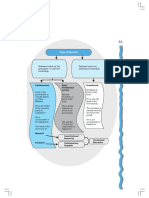
- Bill Becomes Law: Chapter 5: LegislatureDocument1 pageBill Becomes Law: Chapter 5: LegislatureArun MauryaPas encore d'évaluation
- 0-Principle of CryobiologyDocument9 pages0-Principle of CryobiologyArun MauryaPas encore d'évaluation
- Semen and Embryo SexingDocument6 pagesSemen and Embryo SexingArun MauryaPas encore d'évaluation
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5794)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (895)
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (588)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (400)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2259)
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (74)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (345)
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (121)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- Developmental GeneticsDocument18 pagesDevelopmental Geneticsyuyu tuptupPas encore d'évaluation
- Stat151 Final 1Document6 pagesStat151 Final 1INNOVATIVE- Reymar Delos santosPas encore d'évaluation
- Notes From Langman's Medical EmbryologyDocument13 pagesNotes From Langman's Medical EmbryologyJeane Irish Paller Egot92% (12)
- GR 5 ScienceDocument354 pagesGR 5 ScienceGolden SunrisePas encore d'évaluation
- RADBIOLOGY Ievan Dred Balmores OriginalDocument15 pagesRADBIOLOGY Ievan Dred Balmores OriginalMichaela Marie DimasPas encore d'évaluation
- Germ Cell and Fertilization 1Document70 pagesGerm Cell and Fertilization 1Aaryan GuptaPas encore d'évaluation
- Endocrine Regulation of Fish ReproductionDocument10 pagesEndocrine Regulation of Fish ReproductionUmi RehanPas encore d'évaluation
- Animal Biotechnology Test Exam Critical QuestionsDocument73 pagesAnimal Biotechnology Test Exam Critical Questionsbewoketu maru100% (1)
- Budwig Cancer GuideDocument108 pagesBudwig Cancer GuideasoboohbahPas encore d'évaluation
- Origin of Germ Cells.Document12 pagesOrigin of Germ Cells.علیزہ نعیمPas encore d'évaluation
- 22 10 Human Genetics CIBID 2014Document98 pages22 10 Human Genetics CIBID 2014Малиша МанићPas encore d'évaluation
- Carpenter - Pediatric and Adolescent Gynecology 2nd Ed PDFDocument240 pagesCarpenter - Pediatric and Adolescent Gynecology 2nd Ed PDFAngie MirandaPas encore d'évaluation
- Practice Exam 1Document8 pagesPractice Exam 1Tofik MohammedPas encore d'évaluation
- Langman Medical Embryology Made Easy PDFDocument19 pagesLangman Medical Embryology Made Easy PDFAsma SaleemPas encore d'évaluation
- Animal Form and FunctionDocument8 pagesAnimal Form and FunctionPalad , John Carlo BernabePas encore d'évaluation
- Ebook PDF Developmental Biology Tenth 10th Edition PDFDocument41 pagesEbook PDF Developmental Biology Tenth 10th Edition PDFjanet.dicks248100% (42)
- Spermatogenesis: An Overview: Rakesh Sharma and Ashok AgarwalDocument25 pagesSpermatogenesis: An Overview: Rakesh Sharma and Ashok Agarwalumar umarPas encore d'évaluation
- Fertilisation Implantation NotesDocument4 pagesFertilisation Implantation NotesShaun JinPas encore d'évaluation
- Dasar-Dasar Biomolekuler Reproduksi Wanita: Rahmatina B. HermanDocument29 pagesDasar-Dasar Biomolekuler Reproduksi Wanita: Rahmatina B. HermansilmiPas encore d'évaluation
- Oo GenesisDocument20 pagesOo GenesisDinar Yudistira FirdausPas encore d'évaluation
- ANA205 Introduction To Embryology (Lecture 1) Enaibe 2019Document35 pagesANA205 Introduction To Embryology (Lecture 1) Enaibe 2019DOUGLAS KIPRUTOPas encore d'évaluation
- Supp 2017 PDFDocument28 pagesSupp 2017 PDFDesmond BwalyaPas encore d'évaluation
- Regeneration of The Heart: Matthew L. Steinhauser, Richard T. LeeDocument12 pagesRegeneration of The Heart: Matthew L. Steinhauser, Richard T. LeeᄋPas encore d'évaluation
- 2019 Spermatogenesis Biology and Clinical ImplicationsDocument293 pages2019 Spermatogenesis Biology and Clinical ImplicationsPricope Elvis100% (1)
- Basic Embryology Series-1-1Document229 pagesBasic Embryology Series-1-1davidbenedict870Pas encore d'évaluation
- Male and Female Reproductive SystemDocument22 pagesMale and Female Reproductive SystemRhod Vincent Jamolod TayongPas encore d'évaluation
- Larsens Human Embryology 4th Edition Schoenwolf Test BankDocument4 pagesLarsens Human Embryology 4th Edition Schoenwolf Test BankJoshuaKnappkmzpc100% (18)
- Insect Reproduction PDFDocument266 pagesInsect Reproduction PDFJorgePas encore d'évaluation
- Fish Genetics Theory and Practice PDF FormatDocument194 pagesFish Genetics Theory and Practice PDF FormatCristel Mae De GuzmanPas encore d'évaluation
- (Research Progress in Botany) Philip Stewart - Sabine Globig-Reproductive Physiology in Plants-Apple Academic Press (2011)Document321 pages(Research Progress in Botany) Philip Stewart - Sabine Globig-Reproductive Physiology in Plants-Apple Academic Press (2011)Heruko Fatimah100% (1)