Vous aimerez peut-être aussi
- 2010 - UzDocument8 pages2010 - UzCarlPas encore d'évaluation
- Role For Hedgehog Signaling in Cranial-Suture Development and ObesityDocument9 pagesRole For Hedgehog Signaling in Cranial-Suture Development and ObesityDanny Alvarez FocacciPas encore d'évaluation
- Gen-Tieu XuowngDocument21 pagesGen-Tieu XuowngphungPas encore d'évaluation
- Zheng 2013Document6 pagesZheng 2013namiraPas encore d'évaluation
- Novel Human MutationDocument8 pagesNovel Human Mutationnana syahlaPas encore d'évaluation
- ADJALEXU - Volume 44 - Issue 3 - Pages 50-55Document6 pagesADJALEXU - Volume 44 - Issue 3 - Pages 50-55Thien LuPas encore d'évaluation
- 32Document8 pages32Guhan KAPas encore d'évaluation
- Genetics 603: Bamhi G A C Bglii T ADocument8 pagesGenetics 603: Bamhi G A C Bglii T AZenPhiPas encore d'évaluation
- Letter: Genome-Wide Analysis of A Long-Term Evolution Experiment With DrosophilaDocument6 pagesLetter: Genome-Wide Analysis of A Long-Term Evolution Experiment With DrosophilaPeteMossPas encore d'évaluation
- Archives of Oral Biology: RUNX2 in A Chinese Family With Cleidocranial DysplasiaDocument8 pagesArchives of Oral Biology: RUNX2 in A Chinese Family With Cleidocranial DysplasiaDaliz Ü RuizPas encore d'évaluation
- NIH Public Access: Author ManuscriptDocument12 pagesNIH Public Access: Author ManuscriptMaria-Lavinia HoinaruPas encore d'évaluation
- Pang Razi o 2012Document6 pagesPang Razi o 2012Patricio WernerPas encore d'évaluation
- Cone Photoreceptor Structure in Patients With X-Linked Cone Dysfunction and Red-Green Color Vision DeficiencyDocument11 pagesCone Photoreceptor Structure in Patients With X-Linked Cone Dysfunction and Red-Green Color Vision DeficiencyAgnezPas encore d'évaluation
- First HPSE2 Missense Mutation in Urofacial SyndromeDocument5 pagesFirst HPSE2 Missense Mutation in Urofacial SyndromeAmir Akram MasihPas encore d'évaluation
- 1 s2.0 S875632821100980X MainDocument4 pages1 s2.0 S875632821100980X MainKarthik AripakaPas encore d'évaluation
- Letter: Mutations in UBQLN2 Cause Dominant X-Linked Juvenile and Adult-Onset ALS and ALS/dementiaDocument7 pagesLetter: Mutations in UBQLN2 Cause Dominant X-Linked Juvenile and Adult-Onset ALS and ALS/dementiaGarzo ZardofskyPas encore d'évaluation
- DGI TIPE 2 BDocument4 pagesDGI TIPE 2 BYurika HandayaniPas encore d'évaluation
- P63 and IRF6 regulate palate developmentDocument4 pagesP63 and IRF6 regulate palate developmentNadya PurwantyPas encore d'évaluation
- 10 1002@sici1096-9896200003190@4516@@aid-Path5143 0 Co2-J PDFDocument2 pages10 1002@sici1096-9896200003190@4516@@aid-Path5143 0 Co2-J PDFjahdsdjad asffdhsajhajdkPas encore d'évaluation
- De Novo Mutations in YWHAG Cause Early Onset EpilepsyDocument11 pagesDe Novo Mutations in YWHAG Cause Early Onset EpilepsyGiselle Costa Daniel HonoratoPas encore d'évaluation
- jcinvest00095-0208Document7 pagesjcinvest00095-0208bilo47762Pas encore d'évaluation
- DeleciónParcial5pDuplicación_2007Document8 pagesDeleciónParcial5pDuplicación_2007Elizabeth TetelpaPas encore d'évaluation
- Gardner Et Al. - 2016 - Cone Opsins Colour Blindness and Cone Dystrophy Genotype Phenotype CorrelationsDocument4 pagesGardner Et Al. - 2016 - Cone Opsins Colour Blindness and Cone Dystrophy Genotype Phenotype CorrelationsFer FloressPas encore d'évaluation
- Piro Poi Quilo Cito Sis ArticuloDocument6 pagesPiro Poi Quilo Cito Sis ArticuloLaura HernandezPas encore d'évaluation
- JHG CataractDocument3 pagesJHG CataractAyen AlingPas encore d'évaluation
- Syndromes of The First and Second Pharyngeal Arches: A ReviewDocument7 pagesSyndromes of The First and Second Pharyngeal Arches: A ReviewHARITHA H.PPas encore d'évaluation
- ZR 43 2 225Document4 pagesZR 43 2 225Rafay MirzaPas encore d'évaluation
- Brief Report: FOXL2 Mutations in Chinese Families With Blepharophimosis Syndrome (BPES)Document5 pagesBrief Report: FOXL2 Mutations in Chinese Families With Blepharophimosis Syndrome (BPES)flatmajor7Pas encore d'évaluation
- Genome-Wide Association Study Revealed Genomic RegDocument7 pagesGenome-Wide Association Study Revealed Genomic RegGilbert John LacortePas encore d'évaluation
- Resource: A TALEN Genome-Editing System For Generating Human Stem Cell-Based Disease ModelsDocument14 pagesResource: A TALEN Genome-Editing System For Generating Human Stem Cell-Based Disease ModelsfitkaPas encore d'évaluation
- Aku Exon DeletionDocument6 pagesAku Exon DeletionShreyash NaradePas encore d'évaluation
- Hum Mol Genet - 2013-Lee-Hmg ddt022Document16 pagesHum Mol Genet - 2013-Lee-Hmg ddt022api-240674922Pas encore d'évaluation
- mtDNA tRNADocument10 pagesmtDNA tRNAOday MadiPas encore d'évaluation
- Ncomms 10815Document12 pagesNcomms 10815fizzPas encore d'évaluation
- Wu Et Al 2016Document11 pagesWu Et Al 2016MiaPas encore d'évaluation
- Duplication at Xq13.3-q21.1 With Syndromic Intellectual Disability, A Probable Role For The ATRX GeneDocument6 pagesDuplication at Xq13.3-q21.1 With Syndromic Intellectual Disability, A Probable Role For The ATRX GeneEyyup UctepePas encore d'évaluation
- PDE6C Mutations Cause Early-Onset Cone DisordersDocument8 pagesPDE6C Mutations Cause Early-Onset Cone DisordersPablo G. BledtPas encore d'évaluation
- Identification of A Frameshift Mutation in Osterix in A Patient With Recessive Osteogenesis ImperfectaDocument5 pagesIdentification of A Frameshift Mutation in Osterix in A Patient With Recessive Osteogenesis ImperfectaZulia Ahmad BurhaniPas encore d'évaluation
- Deleciones Multiexon en COL1A2Document7 pagesDeleciones Multiexon en COL1A2Alejandra ObandoPas encore d'évaluation
- Cell Sept 2011Document201 pagesCell Sept 2011mekkararameshPas encore d'évaluation
- 2011 Gamba GenetMolResDocument7 pages2011 Gamba GenetMolResEAPPas encore d'évaluation
- Hutchinson-Gilford Progeria Syndrome: Mini ReviewDocument7 pagesHutchinson-Gilford Progeria Syndrome: Mini ReviewBrenda Argomedo CamposPas encore d'évaluation
- Severe Forms of Partial Androgen Insensitivity Syndrome Due To p.L830F Novel Mutation in Androgen Receptor Gene in A Brazilian FamilyDocument6 pagesSevere Forms of Partial Androgen Insensitivity Syndrome Due To p.L830F Novel Mutation in Androgen Receptor Gene in A Brazilian FamilyoktisaripuniPas encore d'évaluation
- Ototoxic Destruction by Co-Administration of Kanamycin and Ethacrynic Acid in RatsDocument9 pagesOtotoxic Destruction by Co-Administration of Kanamycin and Ethacrynic Acid in RatsbeautifulbeastPas encore d'évaluation
- 1 s2.0 S1934590907000677 MainDocument7 pages1 s2.0 S1934590907000677 MainsuperherosavesPas encore d'évaluation
- Article 2Document9 pagesArticle 2hkhanPas encore d'évaluation
- The Ashbya Gossypii Genome As A Tool For Mapping The AncientDocument4 pagesThe Ashbya Gossypii Genome As A Tool For Mapping The AncientJuan RamirezPas encore d'évaluation
- 68th AACC Annual Scientific Meeting Abstract eBookD'Everand68th AACC Annual Scientific Meeting Abstract eBookPas encore d'évaluation
- Leffler 2017Document23 pagesLeffler 2017Dominic AmuzuPas encore d'évaluation
- 6Document5 pages6Iso MogoPas encore d'évaluation
- Amyloid, The Presenilins and Alzheimer's Disease: EviewDocument6 pagesAmyloid, The Presenilins and Alzheimer's Disease: EviewMuhd ShukriPas encore d'évaluation
- Fced GenetikDocument33 pagesFced GenetikyosuanugrahapratamaPas encore d'évaluation
- Funano - Identification of Shared and Unique GeneDocument6 pagesFunano - Identification of Shared and Unique GeneFauzia Latifah SupriyadiPas encore d'évaluation
- Multiple Lipomas Linked To An RB1 Gene Mutation in A Large Pedigree With Low Penetrance RetinoblastomaDocument5 pagesMultiple Lipomas Linked To An RB1 Gene Mutation in A Large Pedigree With Low Penetrance RetinoblastomaMaria Burke-ChaussonPas encore d'évaluation
- Amelogenins: Multi-Functional Proteins in Enamel & Periodontal TissuesDocument13 pagesAmelogenins: Multi-Functional Proteins in Enamel & Periodontal TissuesSABUESO FINANCIEROPas encore d'évaluation
- AMBN2Document18 pagesAMBN2Lisa PrihastariPas encore d'évaluation
- Jahr Et Al., 2005Document6 pagesJahr Et Al., 2005Angelescu LauraPas encore d'évaluation
- LV 2008Document4 pagesLV 2008georgios031Pas encore d'évaluation
- Transcriptome Sequencing of A Large Human Family Identifies The Impact of Rare Noncoding VariantsDocument12 pagesTranscriptome Sequencing of A Large Human Family Identifies The Impact of Rare Noncoding VariantsJacintoOrozcoSamuelPas encore d'évaluation
- Catarata Congenita HereditariaDocument13 pagesCatarata Congenita HereditariaMontze DíazPas encore d'évaluation
- 0 Deec 5269 Aca 990 A 93000000Document11 pages0 Deec 5269 Aca 990 A 93000000Nadya PurwantyPas encore d'évaluation
- 15 147Document5 pages15 147Nadya PurwantyPas encore d'évaluation
- Nadya Purwanty (04121004052) Rkg4Document9 pagesNadya Purwanty (04121004052) Rkg4Nadya PurwantyPas encore d'évaluation
- Comparison of Microleakage of Different Margin Types Around Class V Resin Restorations in Primary TeethDocument6 pagesComparison of Microleakage of Different Margin Types Around Class V Resin Restorations in Primary TeethNadya PurwantyPas encore d'évaluation
- Doxycycline Inhibition of Interleukin-1 in The Corneal EpitheliumDocument14 pagesDoxycycline Inhibition of Interleukin-1 in The Corneal EpitheliumNadya PurwantyPas encore d'évaluation
- Periodontal infectogenomics review examines host genetics and microbiotaDocument6 pagesPeriodontal infectogenomics review examines host genetics and microbiotaNadya PurwantyPas encore d'évaluation
- Microrna-146 Inhibits Pro-Inflammatory Cytokine Secretion Through Il-1 Receptor-Associated Kinase 1 in Human Gingival FibroblastsDocument9 pagesMicrorna-146 Inhibits Pro-Inflammatory Cytokine Secretion Through Il-1 Receptor-Associated Kinase 1 in Human Gingival FibroblastsNadya PurwantyPas encore d'évaluation
- 2012 Bio Matter 0034 RDocument7 pages2012 Bio Matter 0034 RNadya PurwantyPas encore d'évaluation
- 830 840 FGF 2 Induces Proliferation of Human Periodontal Ligament Cells and Maintains DifferentiaDocument11 pages830 840 FGF 2 Induces Proliferation of Human Periodontal Ligament Cells and Maintains DifferentiaNadya PurwantyPas encore d'évaluation
- Simons Pap FinalDocument10 pagesSimons Pap FinalNadya PurwantyPas encore d'évaluation
- Dental Research Journal: Microflora and Periodontal DiseaseDocument5 pagesDental Research Journal: Microflora and Periodontal DiseaseNadya PurwantyPas encore d'évaluation
- Dental Research Journal: Microflora and Periodontal DiseaseDocument5 pagesDental Research Journal: Microflora and Periodontal DiseaseNadya PurwantyPas encore d'évaluation
- Detection of Porphyromonas Gingivalis (FimA) in CoronaryDocument8 pagesDetection of Porphyromonas Gingivalis (FimA) in CoronarySudheer AluruPas encore d'évaluation
- 092 02Document7 pages092 02Nadya PurwantyPas encore d'évaluation
- 110TODENTJDocument6 pages110TODENTJNadya PurwantyPas encore d'évaluation
- CRP and IL-1B Gene Polymorphisms and CRP in Blood in Periodontal DiseaseDocument6 pagesCRP and IL-1B Gene Polymorphisms and CRP in Blood in Periodontal DiseaseNadya PurwantyPas encore d'évaluation
- 6 Journal of Clinical Periodontology 2004 CARD15 Gene Mutations in Adult PeriodontitisDocument4 pages6 Journal of Clinical Periodontology 2004 CARD15 Gene Mutations in Adult PeriodontitisNadya PurwantyPas encore d'évaluation
- 285 FullDocument10 pages285 FullNadya PurwantyPas encore d'évaluation
- 39 1 107aurer enDocument6 pages39 1 107aurer enNadya PurwantyPas encore d'évaluation
- 55 233 1 PBDocument7 pages55 233 1 PBNadya PurwantyPas encore d'évaluation
- 55 233 1 PBDocument7 pages55 233 1 PBNadya PurwantyPas encore d'évaluation
- 10 1 1 321 598Document6 pages10 1 1 321 598Nadya PurwantyPas encore d'évaluation
- 3 D1072 Filiz Acun KayaDocument6 pages3 D1072 Filiz Acun KayaNadya PurwantyPas encore d'évaluation
- 050 23210life110314 346 355Document10 pages050 23210life110314 346 355Nadya PurwantyPas encore d'évaluation
- 030Document13 pages030Nadya PurwantyPas encore d'évaluation
- 14 1 Association Interleukin BayoumiDocument8 pages14 1 Association Interleukin BayoumiNadya PurwantyPas encore d'évaluation
- 14 1 Association Interleukin BayoumiDocument8 pages14 1 Association Interleukin BayoumiNadya PurwantyPas encore d'évaluation
- 02 - Interleukin-1 As A Marker of PeriodontitisDocument3 pages02 - Interleukin-1 As A Marker of PeriodontitisNadya PurwantyPas encore d'évaluation
- The Interleukin-1 Receptor Antagonist (IL-1-Ra) and Soluble Tumor Necrosis Factor Receptor I (STNF RI) in Periodontal DiseaseDocument7 pagesThe Interleukin-1 Receptor Antagonist (IL-1-Ra) and Soluble Tumor Necrosis Factor Receptor I (STNF RI) in Periodontal DiseaseNadya PurwantyPas encore d'évaluation
- Outline For PhotosynthesisDocument6 pagesOutline For Photosynthesiswitzy11Pas encore d'évaluation
- Organic Rankine Cycle (ORC) ExplainedDocument1 pageOrganic Rankine Cycle (ORC) ExplaineddolphinPas encore d'évaluation
- April 2010 One FileDocument296 pagesApril 2010 One FileSaad MotawéaPas encore d'évaluation
- Standby battery product overviewDocument2 pagesStandby battery product overviewHillary McgowanPas encore d'évaluation
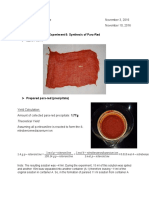
- Synthesis of Para-Red and 1-Iodo-4-Nitrobenzene ExperimentDocument2 pagesSynthesis of Para-Red and 1-Iodo-4-Nitrobenzene ExperimentMaryPas encore d'évaluation
- Carbohydrates OutlineDocument3 pagesCarbohydrates OutlineKalka BoroPas encore d'évaluation
- Thin Layer ChromatographyDocument497 pagesThin Layer ChromatographyFrancimauroPas encore d'évaluation
- Aetheric Battery - Copy #1Document6 pagesAetheric Battery - Copy #1Keresztúri FerencPas encore d'évaluation
- Laboratory Report on Emulsion ExperimentDocument14 pagesLaboratory Report on Emulsion Experimenteizat abasPas encore d'évaluation
- IJCE v5n1p48 enDocument18 pagesIJCE v5n1p48 enAslam HutagiPas encore d'évaluation
- G 010715 RDocument1 pageG 010715 RLiva DesnaPas encore d'évaluation
- U4 - WeldingDocument59 pagesU4 - WeldingAnonymous LM9RaUA4yPas encore d'évaluation
- Syed Sabir - The Recovery of Gold From Secondary Sources-Imperial College Press (2016)Document233 pagesSyed Sabir - The Recovery of Gold From Secondary Sources-Imperial College Press (2016)RODRIGO ARTURO BORJA CALDERÓNPas encore d'évaluation
- Chapter 8 - Synthesis of Nanomaterials - 2009 Ashby - Nanomaterials Nanotechnology An Introduction PDFDocument17 pagesChapter 8 - Synthesis of Nanomaterials - 2009 Ashby - Nanomaterials Nanotechnology An Introduction PDFkaoru9010-1Pas encore d'évaluation
- B001 Arihant PDFDocument12 pagesB001 Arihant PDFmathclubPas encore d'évaluation
- Stowa 2002-11B PDFDocument155 pagesStowa 2002-11B PDFKahl YeongPas encore d'évaluation
- Growing shrimp and tilapia together in indoor aquacultureDocument13 pagesGrowing shrimp and tilapia together in indoor aquaculturetushar patelPas encore d'évaluation
- Inorganic 3Document7 pagesInorganic 3Ephraim PeasahPas encore d'évaluation
- Geotextile BookDocument63 pagesGeotextile BookIsparmoPas encore d'évaluation
- Chapter 1 - Introduction EDMDocument5 pagesChapter 1 - Introduction EDMPraveen RnPas encore d'évaluation
- 50 Years After The Nobel Prize Ziegler Natta Catalysis - Claverie2013Document6 pages50 Years After The Nobel Prize Ziegler Natta Catalysis - Claverie2013Erika AndradePas encore d'évaluation
- Part 2 - AtmosphereDocument21 pagesPart 2 - AtmosphereMarie YabutPas encore d'évaluation
- Gear CalculationDocument58 pagesGear CalculationSachin KubasadPas encore d'évaluation
- Flow of Fluids ReportDocument21 pagesFlow of Fluids ReportalyssaPas encore d'évaluation
- MOF Part 2-1-100Document100 pagesMOF Part 2-1-100Tayyaba BibiPas encore d'évaluation
- Compatibility of Materials With Liquid Oxygen (Impact Sensitivity Threshold and Pass-Fail Techniques)Document12 pagesCompatibility of Materials With Liquid Oxygen (Impact Sensitivity Threshold and Pass-Fail Techniques)Ahmed AlzubaidiPas encore d'évaluation
- Ultrasonic Testing Equipment Manufacturers - Canopus InstrumentsDocument1 pageUltrasonic Testing Equipment Manufacturers - Canopus InstrumentscanopusinstrumentsPas encore d'évaluation
- SM ch1 Mat Meyers 2Document37 pagesSM ch1 Mat Meyers 2infinity_azPas encore d'évaluation
- Reserve Estimate and Development Study of Titas Gas Field PDFDocument109 pagesReserve Estimate and Development Study of Titas Gas Field PDFনাঈমআহমেদPas encore d'évaluation