Vous aimerez peut-être aussi
- Unravelling The Mysteries of Mind and Body Through AbhidhammaDocument157 pagesUnravelling The Mysteries of Mind and Body Through AbhidhammaTanvir100% (3)
- IABU 2012, Buddhist Philosophy and Meditation PracticeDocument488 pagesIABU 2012, Buddhist Philosophy and Meditation Practicekhriniz83% (6)
- Enlightenment by A Single Means, Jackson, 1994Document114 pagesEnlightenment by A Single Means, Jackson, 1994ZacGoPas encore d'évaluation
- Lotus Gospel or Mah 00 Gord RichDocument430 pagesLotus Gospel or Mah 00 Gord RichZacGoPas encore d'évaluation
- General Retreat Reg - Form-Mar.10.2012Document4 pagesGeneral Retreat Reg - Form-Mar.10.2012ZacGoPas encore d'évaluation
- Five Mindfulness Trainings PDFDocument4 pagesFive Mindfulness Trainings PDFZacGoPas encore d'évaluation
- Diamond Sutra v1.9.14 20130105Document40 pagesDiamond Sutra v1.9.14 20130105ZacGoPas encore d'évaluation
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5795)
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (588)
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (74)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (895)
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (400)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (345)
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2259)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (266)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (121)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- Neurology II 6.04 Traumatic Brain Injury Dr. TanDocument10 pagesNeurology II 6.04 Traumatic Brain Injury Dr. TanAbi IgsPas encore d'évaluation
- Genu Valgum Treatment & SurgeryDocument17 pagesGenu Valgum Treatment & SurgeryRanadaPrasadSahaPas encore d'évaluation
- LVS Chronic Pain WsDocument49 pagesLVS Chronic Pain WsdtncorreoPas encore d'évaluation
- Imp-Mediclaim & Accidental Insurance - DataDocument16 pagesImp-Mediclaim & Accidental Insurance - DataAnup BhutadaPas encore d'évaluation
- NAVLE Study Strategies Zuku ReviewDocument60 pagesNAVLE Study Strategies Zuku ReviewMohamed Omar90% (10)
- Analice Mendel: Clinical Doctorate in Occupational Therapy GPA: 4.0Document1 pageAnalice Mendel: Clinical Doctorate in Occupational Therapy GPA: 4.0api-518311936Pas encore d'évaluation
- English Sistem EndocrineDocument9 pagesEnglish Sistem EndocrineNurlaili YaniPas encore d'évaluation
- Complication of Enteral Nutrition PDFDocument3 pagesComplication of Enteral Nutrition PDFIndra WijayaPas encore d'évaluation
- Famador O. Genaldo, MD, RNDocument21 pagesFamador O. Genaldo, MD, RNchance77100% (2)
- Revision 1 English Open LetterDocument9 pagesRevision 1 English Open Letterapi-441660843Pas encore d'évaluation
- Summer Training Guide For Pharmacy Students-1Document132 pagesSummer Training Guide For Pharmacy Students-1Rania T. Sabri100% (2)
- Loneliness Matters. A Theoretical and Empirical Review of Consequences and MechanismsDocument14 pagesLoneliness Matters. A Theoretical and Empirical Review of Consequences and MechanismsIoana UngurianuPas encore d'évaluation
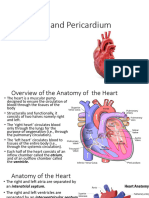
- The Heart and PericardiumDocument40 pagesThe Heart and PericardiumMartha Orendu Oche AttahPas encore d'évaluation
- CHN ExamDocument30 pagesCHN ExamKhei Laqui SN100% (3)
- Cedar Crest College Clinical and Concept Map Worksheet For Nursing 330 PediatricsDocument13 pagesCedar Crest College Clinical and Concept Map Worksheet For Nursing 330 Pediatricsapi-405374041Pas encore d'évaluation
- Practice Guidelines For Obstetric Anesthesia An Updated Report by The American Society of Anesthesiologists Task Force On Obstetric AnesthesiaDocument14 pagesPractice Guidelines For Obstetric Anesthesia An Updated Report by The American Society of Anesthesiologists Task Force On Obstetric AnesthesiaMadalina TalpauPas encore d'évaluation
- 017 Pathology MCQ ACEM Primary GITDocument3 pages017 Pathology MCQ ACEM Primary GITRebeenMustafaPas encore d'évaluation
- SSOPDocument133 pagesSSOPYanuar Rufianti Wongi100% (1)
- Study of The Causes of Mortality in Acute Aluminium Phosphide PoisoningDocument11 pagesStudy of The Causes of Mortality in Acute Aluminium Phosphide PoisoningHany TawfikPas encore d'évaluation
- (EPEC) : Enteropathogenic E. ColiDocument1 page(EPEC) : Enteropathogenic E. ColiBig BearPas encore d'évaluation
- Orthodontic Dental Casts The Case For Routine ArtiDocument8 pagesOrthodontic Dental Casts The Case For Routine ArtiLuminous DentalPas encore d'évaluation
- Final PaperDocument58 pagesFinal PaperAngeline MirasolPas encore d'évaluation
- H1 User Manual (H1E02G10)Document69 pagesH1 User Manual (H1E02G10)Hồng Vân PhạmPas encore d'évaluation
- Lesson Plan NutritionDocument7 pagesLesson Plan NutritionPraveenPas encore d'évaluation
- Acne Herbal TreatmentDocument7 pagesAcne Herbal TreatmentPriyanka Bundhoo100% (1)
- Journal Neuropediatri PDFDocument8 pagesJournal Neuropediatri PDFHalimah PramudiyantiPas encore d'évaluation
- Manila Standard Today - July 21, 2012 IssueDocument16 pagesManila Standard Today - July 21, 2012 IssueManila Standard TodayPas encore d'évaluation
- DR Lie Khie Chen - Management of MDR Infection in Sepsis Jade 2016Document33 pagesDR Lie Khie Chen - Management of MDR Infection in Sepsis Jade 2016Astria PermanaPas encore d'évaluation
- Essay 3. The CNN HeroesDocument3 pagesEssay 3. The CNN HeroesMARIA JOSE CARAVEO GUERREROPas encore d'évaluation