Vous aimerez peut-être aussi
- Falsos CognatosDocument36 pagesFalsos CognatosWillams AlfaiaPas encore d'évaluation
- Curso de Prótese TotalDocument33 pagesCurso de Prótese TotalWillams Alfaia100% (1)
- Processo de Trabalho em Saúde BucalDocument2 pagesProcesso de Trabalho em Saúde BucalWillams AlfaiaPas encore d'évaluation
- Livro FlúorDocument26 pagesLivro FlúorWillams AlfaiaPas encore d'évaluation
- Inglês Aula 05Document114 pagesInglês Aula 05Willams AlfaiaPas encore d'évaluation
- Inglês Aula 04Document128 pagesInglês Aula 04Willams AlfaiaPas encore d'évaluation
- Inglês Aula 06Document115 pagesInglês Aula 06Willams AlfaiaPas encore d'évaluation
- Técnica de Moldagem PDFDocument10 pagesTécnica de Moldagem PDFWillams AlfaiaPas encore d'évaluation
- Doencas Da Polpa e PeriapiceDocument102 pagesDoencas Da Polpa e PeriapiceWillams AlfaiaPas encore d'évaluation
- Inglês Aula 02Document107 pagesInglês Aula 02Willams AlfaiaPas encore d'évaluation
- Inglês Aula 00Document116 pagesInglês Aula 00Willams AlfaiaPas encore d'évaluation
- Inglês Aula 01Document117 pagesInglês Aula 01Willams AlfaiaPas encore d'évaluation
- Inglês Aula 03Document117 pagesInglês Aula 03Willams AlfaiaPas encore d'évaluation
- Implante OdontologicoDocument6 pagesImplante OdontologicoWillams AlfaiaPas encore d'évaluation
- Tipos de Roscas de ImplantesDocument10 pagesTipos de Roscas de ImplantesWillams AlfaiaPas encore d'évaluation
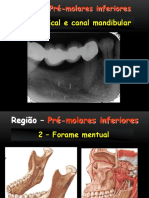
- Pré e Molares InferioresDocument17 pagesPré e Molares InferioreswalfaiaPas encore d'évaluation
- Resumo Patologia OralDocument145 pagesResumo Patologia OralWillams AlfaiaPas encore d'évaluation
- Nomenclatura e Partes Constituintes Das CavidadesDocument4 pagesNomenclatura e Partes Constituintes Das CavidadesWillams AlfaiaPas encore d'évaluation
- Bíblia Revelada Di Nelson - Carta de Judas PDFDocument10 pagesBíblia Revelada Di Nelson - Carta de Judas PDFGenildo FreitasPas encore d'évaluation
- Endodontia Pré-Clinica PDFDocument101 pagesEndodontia Pré-Clinica PDFDanielle SilvaPas encore d'évaluation
- Exegese de Textos Difíceis Da BíbliaDocument187 pagesExegese de Textos Difíceis Da BíbliaMaurício Dâmaso100% (1)
- Passos Clínicos para Confecção de Uma PPRDocument2 pagesPassos Clínicos para Confecção de Uma PPRWillams AlfaiaPas encore d'évaluation
- Carie DentariaDocument63 pagesCarie DentariaWillams AlfaiaPas encore d'évaluation
- Exegese de Textos Difíceis Da BíbliaDocument187 pagesExegese de Textos Difíceis Da BíbliaMaurício Dâmaso100% (1)
- Nomenclatura e Classificação Das Cavidades DentáriasDocument2 pagesNomenclatura e Classificação Das Cavidades DentáriasWillams AlfaiaPas encore d'évaluation
- Alterações Pulpares e Periapicais Prof CarolinaDocument48 pagesAlterações Pulpares e Periapicais Prof CarolinaWillams AlfaiaPas encore d'évaluation
- En Do Don Ti ADocument35 pagesEn Do Don Ti AKyze QuintelaPas encore d'évaluation
- En Do Don Ti ADocument35 pagesEn Do Don Ti AKyze QuintelaPas encore d'évaluation
- Super Interessante A Origem Da BibliaDocument9 pagesSuper Interessante A Origem Da BibliaWellington LiraPas encore d'évaluation
- Resumo de Grampos para IsolamentoDocument3 pagesResumo de Grampos para IsolamentoWillams AlfaiaPas encore d'évaluation
- Alterações Fisiológicas Do EnvelhecimentoDocument5 pagesAlterações Fisiológicas Do EnvelhecimentoAndersonCarvalho100% (1)
- A Ginástica em Sua Forma Mais ÍntimaDocument4 pagesA Ginástica em Sua Forma Mais ÍntimaRubia Oliveira Oliveira100% (1)
- Ficha Técnica para Terapeutas - Herpes SimplesDocument1 pageFicha Técnica para Terapeutas - Herpes SimplesEspelhodAlmaPas encore d'évaluation
- Como Funciona o Cérebro de Uma Pessoa ApaixonadaDocument23 pagesComo Funciona o Cérebro de Uma Pessoa ApaixonadaNatália BrasilPas encore d'évaluation
- PoliglotaDocument3 pagesPoliglotaRita BatistaPas encore d'évaluation
- Buracos Da Base Do CrânioDocument3 pagesBuracos Da Base Do CrânioInês FerreiraPas encore d'évaluation
- AULA DE BIOELETROGÊNESE 2014 AlunosDocument53 pagesAULA DE BIOELETROGÊNESE 2014 AlunosLhana Lorena Melo AtanasioPas encore d'évaluation
- Apostila de MicroagulhamentoDocument47 pagesApostila de Microagulhamentodanielle100% (13)
- Ento OrdensDocument108 pagesEnto OrdensKim Kishi Sena100% (1)
- Curativo e Tipos de CurativosDocument5 pagesCurativo e Tipos de Curativosafsilva_enfermagem5887100% (1)
- Aula DoaçãoDocument70 pagesAula DoaçãoRicardo Alessandro Teixeira GonsagaPas encore d'évaluation
- EdemaDocument2 pagesEdemaMauro Neto100% (1)
- Guia ESDocument152 pagesGuia ESPablo RibeiroPas encore d'évaluation
- AULA 8 - HanseníaseDocument80 pagesAULA 8 - HanseníaseAvhaPaixão100% (3)
- Lygia Fagundes Telles - Biruta - RevistoDocument9 pagesLygia Fagundes Telles - Biruta - RevistoGian SilvaPas encore d'évaluation
- Cartilha Os InsetosDocument19 pagesCartilha Os InsetosGeferson Fernando MetzPas encore d'évaluation
- Caixa TorácicaDocument7 pagesCaixa TorácicaAlline DO Valle100% (1)
- 1° SIMULADO CADERNO DE QUESTÕES - Compressed PDFDocument13 pages1° SIMULADO CADERNO DE QUESTÕES - Compressed PDFCésar MascarenhaPas encore d'évaluation
- Síndrome Do Definhamento Do Neonato - Saiba Tudo Sobre Síndrome Do Definhamento Do NeonatoDocument1 pageSíndrome Do Definhamento Do Neonato - Saiba Tudo Sobre Síndrome Do Definhamento Do NeonatowendyelagoPas encore d'évaluation
- Evolução Do Grau de Incapacidade Física de Um Paciente Com Hanseníase - Relato de CasoDocument1 pageEvolução Do Grau de Incapacidade Física de Um Paciente Com Hanseníase - Relato de CasoSuzane Ketlyn MartelloPas encore d'évaluation
- Apostila Introducao A Saude BucalDocument100 pagesApostila Introducao A Saude BucalCintya Silva100% (2)
- LIVRO DE NATUROLOGIA CIÊNCIA E VIDA - Parte IDocument102 pagesLIVRO DE NATUROLOGIA CIÊNCIA E VIDA - Parte IAPH - Associação Portuguesa de Homeopatia (repositório 2008-15)86% (7)
- 1º Teste 6º10Document9 pages1º Teste 6º10Rosa Teixeira50% (2)
- Veja Como É o Rottsky, o Cão Mistura de RottweileDocument1 pageVeja Como É o Rottsky, o Cão Mistura de RottweileRicardo Moreno RachelPas encore d'évaluation
- Tecido ConjuntivoDocument16 pagesTecido ConjuntivoGabriela BrazilPas encore d'évaluation
- Apostila BambuterapiaDocument8 pagesApostila BambuterapiaAdriana Queiroz75% (4)
- AlimentaçãoDocument39 pagesAlimentaçãoraubersg8015Pas encore d'évaluation
- Curso Básico - A Era Dos Cristais 4.0Document22 pagesCurso Básico - A Era Dos Cristais 4.0sanandrasan100% (1)
- Manual Tecnico Hiv 27 11 2018 Web PDFDocument148 pagesManual Tecnico Hiv 27 11 2018 Web PDFRosana Moraes Da SilvaPas encore d'évaluation
- Resumo Anatomia MuscularDocument4 pagesResumo Anatomia MuscularPedro Augusto PerryPas encore d'évaluation