Vous aimerez peut-être aussi
- 50 Great Myths of Human Evolution: Understanding Misconceptions about Our OriginsD'Everand50 Great Myths of Human Evolution: Understanding Misconceptions about Our OriginsPas encore d'évaluation
- Mouse Genetic PortraitDocument24 pagesMouse Genetic PortraitbelovedaffectionPas encore d'évaluation
- Group 6-3ebio: Assoc Prof. Loida R. Medina, PHD, RMT, RmicroDocument4 pagesGroup 6-3ebio: Assoc Prof. Loida R. Medina, PHD, RMT, RmicroAnne Olfato ParojinogPas encore d'évaluation
- 2010 (Quin Et Al.) A Human Gut Microbial Gene Catalogue Established by Metagenomic SequencingDocument9 pages2010 (Quin Et Al.) A Human Gut Microbial Gene Catalogue Established by Metagenomic SequencingDaniel AlonsoPas encore d'évaluation
- 2017 Nature21674-Neanderthal Behaviour, Diet, and DiseaseDocument16 pages2017 Nature21674-Neanderthal Behaviour, Diet, and DiseaseBernat BertomeuPas encore d'évaluation
- AiG - Genetics - DNA SimilaritiesDocument102 pagesAiG - Genetics - DNA Similaritiesst iPas encore d'évaluation
- Backhed Etal 05 Bac MutualismDocument6 pagesBackhed Etal 05 Bac Mutualismu77Pas encore d'évaluation
- Evolutionary Patterns of RNA-Based Duplication in Non-Mammalian ChordatesDocument9 pagesEvolutionary Patterns of RNA-Based Duplication in Non-Mammalian ChordatesFrontiersPas encore d'évaluation
- The Pineapple Genome and The Evolution of CAM PhotosynthesisDocument11 pagesThe Pineapple Genome and The Evolution of CAM Photosynthesisomprakash2592Pas encore d'évaluation
- C Elegans DissertationDocument7 pagesC Elegans DissertationBuySchoolPapersOnlineSingapore100% (1)
- Genome Res.-2013-Alföldi-1063-8Document7 pagesGenome Res.-2013-Alföldi-1063-8Yamile A Rodríguez RiascosPas encore d'évaluation
- HUMAN GENOME PROJECT DOCUMENTDocument21 pagesHUMAN GENOME PROJECT DOCUMENTSanjay KumarPas encore d'évaluation
- Chromosome-Scale Genome Assembly of Lepus OiostoluDocument7 pagesChromosome-Scale Genome Assembly of Lepus OiostoluMuhammad UsamaPas encore d'évaluation
- tmp1453 TMPDocument6 pagestmp1453 TMPFrontiersPas encore d'évaluation
- Black & White Deliver Card (1) - WPS OfficeDocument28 pagesBlack & White Deliver Card (1) - WPS Officelinkgogo69Pas encore d'évaluation
- The Genetic Basis of Tail-Loss Evolution in Humans and ApesDocument33 pagesThe Genetic Basis of Tail-Loss Evolution in Humans and ApesROCIO CERVANTES HERNANDEZPas encore d'évaluation
- Human Pathways in Animal Models-5aDocument13 pagesHuman Pathways in Animal Models-5aElvina DhamayantiPas encore d'évaluation
- Marshall and Berbee 2010 MBEDocument13 pagesMarshall and Berbee 2010 MBEWyth MarshallPas encore d'évaluation
- Letter: Genome-Wide Analysis of A Long-Term Evolution Experiment With DrosophilaDocument6 pagesLetter: Genome-Wide Analysis of A Long-Term Evolution Experiment With DrosophilaPeteMossPas encore d'évaluation
- Adams Science 1999Document11 pagesAdams Science 1999vnq6gd6qbxPas encore d'évaluation
- Biol 230W Lab ReportDocument8 pagesBiol 230W Lab Reportcsh5213Pas encore d'évaluation
- The Complete Mitochondrial Genome of An 11,450-Year-Old Aurochsen (Bos Primigenius) From Central ItalyDocument10 pagesThe Complete Mitochondrial Genome of An 11,450-Year-Old Aurochsen (Bos Primigenius) From Central ItalyradulusPas encore d'évaluation
- Evolution Evidence ExplainedDocument13 pagesEvolution Evidence ExplainedJohn Bernard VeloriaPas encore d'évaluation
- Sivavel BioDocument14 pagesSivavel BioVigneshPas encore d'évaluation
- Et Al. - On The Origin of LeprosyDocument15 pagesEt Al. - On The Origin of LeprosyRoberto SanchezPas encore d'évaluation
- FOXP2 and NeanderthalsDocument12 pagesFOXP2 and NeanderthalsSxoutPas encore d'évaluation
- T H G P: HE Uman Enome RojectDocument28 pagesT H G P: HE Uman Enome RojectUpendra Sharma Sulibele100% (1)
- Ancient DNA research reveals human evolutionDocument6 pagesAncient DNA research reveals human evolutionAlfiansyah PrasetyoPas encore d'évaluation
- BIOLOGY Class 12 ProjectDocument15 pagesBIOLOGY Class 12 ProjectNaveen Doss75% (185)
- Exome Capture From SalivaDocument17 pagesExome Capture From SalivaJobin John PR15BI1001Pas encore d'évaluation
- PNAS 2010 Huff 2147 52Document6 pagesPNAS 2010 Huff 2147 52Vlad PredaPas encore d'évaluation
- The Molecular Basis of Human Brain Evolution: A Framework for Indirect EvidenceDocument9 pagesThe Molecular Basis of Human Brain Evolution: A Framework for Indirect EvidenceShafat HasanPas encore d'évaluation
- Los Tunicados y Los No Cefalocordados Son Los Parientes Vivos Más Cercanos de Los Vertebrados. 2008Document49 pagesLos Tunicados y Los No Cefalocordados Son Los Parientes Vivos Más Cercanos de Los Vertebrados. 2008BRAYANPas encore d'évaluation
- 3 Model GenomesDocument4 pages3 Model GenomesParisha SinghPas encore d'évaluation
- 2012 - Human Brain Evolution From Gene Discovery To Phenotype DiscoveryDocument8 pages2012 - Human Brain Evolution From Gene Discovery To Phenotype DiscoveryMauricio RíosPas encore d'évaluation
- Biology Methods and General Paper: B. Sc. Degree Examinations 2010Document4 pagesBiology Methods and General Paper: B. Sc. Degree Examinations 2010gggtdyfhuuoPas encore d'évaluation
- 1.1.3.16. Humans Report On Their Own Peculiarities: 22 Chapter 1. I. Introduction To The CellDocument1 page1.1.3.16. Humans Report On Their Own Peculiarities: 22 Chapter 1. I. Introduction To The CellAnna YepPas encore d'évaluation
- Minireview: Ancient DNA and The Polymerase Chain ReactionDocument4 pagesMinireview: Ancient DNA and The Polymerase Chain ReactiontaufiqPas encore d'évaluation
- Human GenomeDocument8 pagesHuman GenomeKannanSubramanianPas encore d'évaluation
- Man and Animals Are 96 Percent The SameDocument86 pagesMan and Animals Are 96 Percent The Samemaqsood hasniPas encore d'évaluation
- Genome Mapping and Genomics in AnimalsDocument289 pagesGenome Mapping and Genomics in AnimalsCarmenPas encore d'évaluation
- The Human Genome ProjectsDocument4 pagesThe Human Genome ProjectsMurugesan PalrajPas encore d'évaluation
- Kolder 2016 A Full Body Transcriptome and ProteDocument12 pagesKolder 2016 A Full Body Transcriptome and ProteAndreea ȘerbanPas encore d'évaluation
- Marine Thesis PDFDocument6 pagesMarine Thesis PDFRaquel Pellicier100% (1)
- Comp Transcript Between Zebrafsh and MammalsDocument5 pagesComp Transcript Between Zebrafsh and MammalsRita RodriguesPas encore d'évaluation
- The Biology of ColorDocument24 pagesThe Biology of ColorMădălina MarincaşPas encore d'évaluation
- Biology Ip HGPDocument10 pagesBiology Ip HGPDipraPas encore d'évaluation
- Eight Thousand Years of Natural Selection in EuropeDocument22 pagesEight Thousand Years of Natural Selection in EuropeRodrick JohnsonPas encore d'évaluation
- Biology Investigatory ProjectDocument44 pagesBiology Investigatory ProjectVaishnavi AvisagaddaPas encore d'évaluation
- Gen 1801619Document10 pagesGen 1801619Luis HernanPas encore d'évaluation
- Diversityscanner: Robotic Discovery of Small Invertebrates With Machine Learning MethodsDocument15 pagesDiversityscanner: Robotic Discovery of Small Invertebrates With Machine Learning MethodsSomesh ReddyPas encore d'évaluation
- Analysis of Streptococcal CRISPRs From Human Saliva Reveals Substantial Sequence Diversity Within and Between Subjects Over TimeDocument12 pagesAnalysis of Streptococcal CRISPRs From Human Saliva Reveals Substantial Sequence Diversity Within and Between Subjects Over Timeabcder1234Pas encore d'évaluation
- Similarities Between Men and AnimalsDocument154 pagesSimilarities Between Men and Animalsmaqsood hasniPas encore d'évaluation
- Test Bank For Molecular Biology of The Cell 5th Edition Bruce AlbertsDocument5 pagesTest Bank For Molecular Biology of The Cell 5th Edition Bruce AlbertsJames Jones100% (6)
- Initial Impact of The Sequencing of The Human GenomeDocument11 pagesInitial Impact of The Sequencing of The Human GenomeMariano PerezPas encore d'évaluation
- FOXP2 Targets Show Evidence of Positive Selection in European PopulationsDocument11 pagesFOXP2 Targets Show Evidence of Positive Selection in European PopulationsTefnut NixPas encore d'évaluation
- Splicing AlternativoDocument8 pagesSplicing AlternativoJon2170Pas encore d'évaluation
- HUMAN GENOME PROJECT: KEY MILESTONES AND FUTURE CHALLENGESDocument23 pagesHUMAN GENOME PROJECT: KEY MILESTONES AND FUTURE CHALLENGESNishant ChandavarkarPas encore d'évaluation
- Lost Nirvana Kedar KelkarDocument254 pagesLost Nirvana Kedar KelkarKedar KelkarPas encore d'évaluation
- SportzVillage Academies OverviewDocument19 pagesSportzVillage Academies OverviewKedar KelkarPas encore d'évaluation
- The Archetypal Triad PDFDocument3 pagesThe Archetypal Triad PDFKedar KelkarPas encore d'évaluation
- Man Must Awaken From His Mental Prison Before Emerging Into His True FormDocument20 pagesMan Must Awaken From His Mental Prison Before Emerging Into His True FormKedar KelkarPas encore d'évaluation
- Imaginary TimeDocument2 pagesImaginary TimeKedar KelkarPas encore d'évaluation
- Godbole Book On Taj PDFDocument118 pagesGodbole Book On Taj PDFpadeepPas encore d'évaluation
- Hypnotherapy Scripts 6 Steve G Jones Ebook PDFDocument3 pagesHypnotherapy Scripts 6 Steve G Jones Ebook PDFKedar KelkarPas encore d'évaluation
- The Hyparxis of The Dramatic Universe PDFDocument7 pagesThe Hyparxis of The Dramatic Universe PDFKedar KelkarPas encore d'évaluation
- Structure of The Present MomentDocument4 pagesStructure of The Present MomentKedar KelkarPas encore d'évaluation
- Mysterious Metaphysics PDFDocument2 pagesMysterious Metaphysics PDFKedar KelkarPas encore d'évaluation
- Man Must Awaken From His Mental Prison Before Emerging Into His True FormDocument20 pagesMan Must Awaken From His Mental Prison Before Emerging Into His True FormKedar KelkarPas encore d'évaluation
- Hypnotherapy Scripts 6 Steve G Jones Ebook PDFDocument66 pagesHypnotherapy Scripts 6 Steve G Jones Ebook PDFjohannes2212100% (10)
- Fowler KennedyDocument15 pagesFowler Kennedypankajgupta86Pas encore d'évaluation
- Alfred P Sloan My Years With General MotorsDocument12 pagesAlfred P Sloan My Years With General MotorsKedar KelkarPas encore d'évaluation
- ACL Reconstruction ProtocolDocument23 pagesACL Reconstruction ProtocolRoberto Tommasini100% (2)
- Yogastha - Sameer Kulkarni PDFDocument17 pagesYogastha - Sameer Kulkarni PDFDileep KumarPas encore d'évaluation
- ODonnell, Gerald - Remote InfluencingDocument27 pagesODonnell, Gerald - Remote InfluencingJeremy CastilloPas encore d'évaluation
- ACL Reconstruction Post Operative Rehabilitation Protocol: Phase I: 1 - 14 Days GoalsDocument5 pagesACL Reconstruction Post Operative Rehabilitation Protocol: Phase I: 1 - 14 Days GoalsFábio SantosPas encore d'évaluation
- Sentence FragmentsDocument4 pagesSentence Fragmentsmacaroanele100% (1)
- The Art of MemeticsDocument239 pagesThe Art of MemeticsWes Unruh97% (34)
- Functional Movement Screen Score Sheet: Test Raw Score Final Score CommentsDocument1 pageFunctional Movement Screen Score Sheet: Test Raw Score Final Score CommentsdiskraPas encore d'évaluation
- Electronic Balance Board ManualDocument40 pagesElectronic Balance Board ManualKedar KelkarPas encore d'évaluation
- Hazrat BabajanDocument12 pagesHazrat BabajanKedar KelkarPas encore d'évaluation
- FMS Scoring CriteriaDocument7 pagesFMS Scoring CriteriaTrainer Aong100% (1)
- NLP Secrets PDFDocument64 pagesNLP Secrets PDFchanayirePas encore d'évaluation
- The Diary of A Modern Alchemist by J H Reyner 1974 PDFDocument97 pagesThe Diary of A Modern Alchemist by J H Reyner 1974 PDFKedar Kelkar100% (3)
- Postural AssessmentDocument2 pagesPostural AssessmentKedar KelkarPas encore d'évaluation
- Timothy Gallwey Meta States and Inner GameDocument12 pagesTimothy Gallwey Meta States and Inner GameGil Eng Mesquita100% (1)
- (Mysticism) Goettmann, Alphonse - Dailog On The Path of Initiation - The Life and Thought of Karlfried Graf DurckheimDocument74 pages(Mysticism) Goettmann, Alphonse - Dailog On The Path of Initiation - The Life and Thought of Karlfried Graf DurckheimAvadon4egPas encore d'évaluation
- Hidden Root PDFDocument6 pagesHidden Root PDFKedar KelkarPas encore d'évaluation
- Ubiquitin Family Modifiers and The Proteasome Reviews and Protocols by Alexander Varshavsky (Auth.), R. Jürgen Dohmen, Martin Scheffner (Eds.)Document660 pagesUbiquitin Family Modifiers and The Proteasome Reviews and Protocols by Alexander Varshavsky (Auth.), R. Jürgen Dohmen, Martin Scheffner (Eds.)Andonis AngelovPas encore d'évaluation
- Faculty of ScienceDocument50 pagesFaculty of SciencesindhujanaPas encore d'évaluation
- Chemistry Biology Exam 2 With Solution PDFDocument26 pagesChemistry Biology Exam 2 With Solution PDFDawlat SalamaPas encore d'évaluation
- New Scientist 06 Aug 2022Document62 pagesNew Scientist 06 Aug 2022BSPas encore d'évaluation
- Recombinant Therapeutic Protein Production in Cultivated Mammalian Cells: Current Status and Future ProspectsDocument6 pagesRecombinant Therapeutic Protein Production in Cultivated Mammalian Cells: Current Status and Future ProspectsGryseldaPas encore d'évaluation
- IgY Proteome Partitioning SolutionsDocument12 pagesIgY Proteome Partitioning SolutionsmfsmarcilioPas encore d'évaluation
- You Are What You EatDocument3 pagesYou Are What You EatOlavo BenevenutoPas encore d'évaluation
- Biology: Organic CompoundsDocument28 pagesBiology: Organic CompoundsMockinjayPas encore d'évaluation
- Gene To Protein, Part 1Document5 pagesGene To Protein, Part 1shiyiPas encore d'évaluation
- Bio Module 1Document43 pagesBio Module 1XdxdPas encore d'évaluation
- Balanced DietDocument13 pagesBalanced DietOffice AccountPas encore d'évaluation
- Q3 G11 Physical Science Module 7Document19 pagesQ3 G11 Physical Science Module 7Lebz RicaramPas encore d'évaluation
- Molecular Cell Biology 1 Basics of Molecular BiologyDocument80 pagesMolecular Cell Biology 1 Basics of Molecular BiologyIana Mira100% (1)
- 01 Intact CaseinDocument38 pages01 Intact CaseinPat FerrerPas encore d'évaluation
- Diagnostic BioSystems CatalogDocument116 pagesDiagnostic BioSystems CatalogCris StefanPas encore d'évaluation
- APSCHE Syllabus for BiochemistryDocument21 pagesAPSCHE Syllabus for BiochemistryS V S VardhanPas encore d'évaluation
- Yersinia Pestis V Protein Epitopes Recognized by CD4 T CellsDocument8 pagesYersinia Pestis V Protein Epitopes Recognized by CD4 T CellsHerdi ArdianaPas encore d'évaluation
- Integrated Laboratory Class 6Document7 pagesIntegrated Laboratory Class 6Neils ArenósPas encore d'évaluation
- 2019 Book PrincipleOfNursingInOncologyDocument454 pages2019 Book PrincipleOfNursingInOncologyMarPas encore d'évaluation
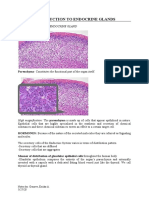
- Introduction To Endocrine GlandsDocument6 pagesIntroduction To Endocrine GlandsEricka GenovePas encore d'évaluation
- Harmening Chapter Summary and Questions CompilationDocument63 pagesHarmening Chapter Summary and Questions CompilationShaira Rae BillenaPas encore d'évaluation
- Biotech q1 Mod1Document32 pagesBiotech q1 Mod1Jem Culianan0% (2)
- Functions of BloodDocument8 pagesFunctions of BloodAndrés Lasso SPas encore d'évaluation
- Topic 1 MCDocument45 pagesTopic 1 MCfrostized001Pas encore d'évaluation
- Full Download Beckers World of The Cell 9th Edition Hardin Solutions ManualDocument35 pagesFull Download Beckers World of The Cell 9th Edition Hardin Solutions Manualsmallmanclaude100% (38)
- Molecular Cell Biology 7Th Edition Full ChapterDocument41 pagesMolecular Cell Biology 7Th Edition Full Chapterjoseph.backman295100% (21)
- IT Bio F4 Topical Test 4 (BL)Document8 pagesIT Bio F4 Topical Test 4 (BL)Ismaliza IshakPas encore d'évaluation
- Physical Science Grade 11 - St. Lorenzo Module 2 - : Saint Louis School of Pacdal, IncDocument10 pagesPhysical Science Grade 11 - St. Lorenzo Module 2 - : Saint Louis School of Pacdal, IncNo namePas encore d'évaluation
- Handbook of Computational Chemistry - Leszczynski P 1110Document3 pagesHandbook of Computational Chemistry - Leszczynski P 1110leofloatingPas encore d'évaluation
- Chemical Properties and Nutritive Value of Milk LectureDocument55 pagesChemical Properties and Nutritive Value of Milk LectureUmesh PoudelPas encore d'évaluation