Vous aimerez peut-être aussi
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (895)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5794)
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (399)
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (73)
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (588)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (344)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2259)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (120)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- Gastrointestinal SystemDocument109 pagesGastrointestinal SystemRaju Mehta100% (1)
- FundusDocument10 pagesFundusKristin NataliaPas encore d'évaluation
- Block Hepatobiliary SystemDocument29 pagesBlock Hepatobiliary SystemKevin KusumanPas encore d'évaluation
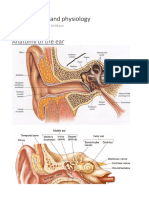
- ENT Anatomy and PhysiologyDocument32 pagesENT Anatomy and Physiologyoxford_commaPas encore d'évaluation
- Short Tricks Biology 11thsDocument36 pagesShort Tricks Biology 11thskomatipallis615Pas encore d'évaluation
- Diabetes MellitusDocument18 pagesDiabetes Mellitusrajirajesh100% (1)
- 3 Tissues in The Human Body 22864Document11 pages3 Tissues in The Human Body 22864Musharaf RehmanPas encore d'évaluation
- Hypothalamus Pituitary Glands Adrenal CortexDocument2 pagesHypothalamus Pituitary Glands Adrenal CortexEmPas encore d'évaluation
- 1757 1626 1 333Document5 pages1757 1626 1 333Ade CandraPas encore d'évaluation
- Science5 q2 Mod1 ThePartsOfTheReproductiveSystemAndTheirFunctions v2Document24 pagesScience5 q2 Mod1 ThePartsOfTheReproductiveSystemAndTheirFunctions v2Shiela AsiloPas encore d'évaluation
- General Physiology Control Systems of Homeostasis: By: Dr. Hamza Al-TradDocument22 pagesGeneral Physiology Control Systems of Homeostasis: By: Dr. Hamza Al-TradibrahimPas encore d'évaluation
- Thyroid Status Examination OSCE GuideDocument20 pagesThyroid Status Examination OSCE GuideemmaPas encore d'évaluation
- K10 Kuliah NefrourologiDocument170 pagesK10 Kuliah NefrourologimarinanananaPas encore d'évaluation
- 1 6 Regulation of Blood Glucose PDFDocument3 pages1 6 Regulation of Blood Glucose PDFtiaraPas encore d'évaluation
- Icopim Vol 2Document169 pagesIcopim Vol 2DannKusumaPas encore d'évaluation
- Themenstrualcycle 120606211927 Phpapp02Document16 pagesThemenstrualcycle 120606211927 Phpapp02Ludy LynPas encore d'évaluation
- Clinical Oriented Anatomy of Urinary SystemDocument81 pagesClinical Oriented Anatomy of Urinary SystemRizcky Naldy Eka Putra100% (1)
- Mote - 'Hugh'Document18 pagesMote - 'Hugh'Melissa R.Pas encore d'évaluation
- Medical Surgical Nursing OrthopedicDocument22 pagesMedical Surgical Nursing Orthopedicroger80% (5)
- KOYANO Et Al-2012-Journal of Oral Rehabilitation PDFDocument9 pagesKOYANO Et Al-2012-Journal of Oral Rehabilitation PDFKathy Cuevas HinojosaPas encore d'évaluation
- Radiology in Pediatric Dentistry 2Document44 pagesRadiology in Pediatric Dentistry 2Aima Cuba100% (1)
- Evaluation of Length Intestine Ratio With Long Body of FishDocument10 pagesEvaluation of Length Intestine Ratio With Long Body of FishPuri RahmaPas encore d'évaluation
- Anatomy of Urinary Bladder and UrethraDocument16 pagesAnatomy of Urinary Bladder and UrethraAmalina ZolkefleePas encore d'évaluation
- Science 10 Q3 - M2Document14 pagesScience 10 Q3 - M2Avha CortesPas encore d'évaluation
- Kents New RemediesDocument168 pagesKents New RemediesluckyPas encore d'évaluation
- Human Reproduction PDFDocument35 pagesHuman Reproduction PDFsqhaa100% (4)
- Endocrine System ChartDocument2 pagesEndocrine System ChartValeria Guadalupe Ramírez Moctezuma100% (2)
- Blood Distribution During Rest and ExerciseDocument2 pagesBlood Distribution During Rest and ExercisePannir SelvaPas encore d'évaluation
- (PED) T.10.2 - Pediatric Endocrinology (Part II) PDFDocument7 pages(PED) T.10.2 - Pediatric Endocrinology (Part II) PDFLiberty AgcaoiliPas encore d'évaluation
- The Heart Has Several Pacemakers Known As Autonomic FociDocument1 pageThe Heart Has Several Pacemakers Known As Autonomic FociAnonymous mLYupGyNPas encore d'évaluation