Vous aimerez peut-être aussi
- Dray e Legendre, 2008 - EcologyDocument21 pagesDray e Legendre, 2008 - EcologyThyago VidovixPas encore d'évaluation
- Dna AlienDocument15 pagesDna AlienThyago VidovixPas encore d'évaluation
- Toward A Smarter EnterpriseDocument29 pagesToward A Smarter EnterpriseThyago VidovixPas encore d'évaluation
- Brookfield, 2000 - What DeterminesDocument2 pagesBrookfield, 2000 - What DeterminesThyago VidovixPas encore d'évaluation
- J. Nutr.-1999-Morita-1731-6Document6 pagesJ. Nutr.-1999-Morita-1731-6Thyago VidovixPas encore d'évaluation
- Single Molecule AnalysisDocument5 pagesSingle Molecule AnalysisThyago VidovixPas encore d'évaluation
- 07Document5 pages07Thyago VidovixPas encore d'évaluation
- Guide Chapter 1Document6 pagesGuide Chapter 1Thyago VidovixPas encore d'évaluation
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (399)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (894)
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (265)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (73)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (119)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- Cell Membrane and Transport ColoringDocument3 pagesCell Membrane and Transport ColoringTeresa GonzPas encore d'évaluation
- MYCOVIRODocument11 pagesMYCOVIROPau SorianoPas encore d'évaluation
- Non-symbiotic Nitrogen FixersDocument21 pagesNon-symbiotic Nitrogen Fixersrajiv pathakPas encore d'évaluation
- Management of PPHDocument1 pageManagement of PPH098 U.KARTHIK SARAVANA KANTHPas encore d'évaluation
- WCH03 01 Que 20180124Document16 pagesWCH03 01 Que 20180124Rameez Mazhar SiddiqiPas encore d'évaluation
- 8 Cell - The Unit of Life-NotesDocument6 pages8 Cell - The Unit of Life-NotesBhavanya RavichandrenPas encore d'évaluation
- Brigitte King - InformativeDocument2 pagesBrigitte King - Informativeapi-262025468Pas encore d'évaluation
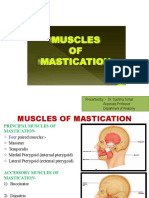
- Presented By:-Dr. Sushma Tomar Associate Professor Department of AnatomyDocument20 pagesPresented By:-Dr. Sushma Tomar Associate Professor Department of AnatomyCristine EchavePas encore d'évaluation
- Parts, Functions and Hormones in Male and Female Reproductive SystemsDocument32 pagesParts, Functions and Hormones in Male and Female Reproductive SystemsZephie100% (1)
- Vanders Human Physiology The Mechanisms of Body Function 15th Edition Widmaier Test BankDocument29 pagesVanders Human Physiology The Mechanisms of Body Function 15th Edition Widmaier Test BankDavidWardrcobi100% (15)
- Citric Acid Production Stirred TankDocument9 pagesCitric Acid Production Stirred TankKarliiux MedinaPas encore d'évaluation
- 421 Reproduction Developmental Toxicity Screening TestDocument10 pages421 Reproduction Developmental Toxicity Screening TestAshishPas encore d'évaluation
- Ancient Healing Rods Reveal Secrets of PharaohsDocument28 pagesAncient Healing Rods Reveal Secrets of PharaohsMiguel Angelum100% (4)
- HHS Public Access: Structure and Function of The Human Skin MicrobiomeDocument20 pagesHHS Public Access: Structure and Function of The Human Skin MicrobiomeEugeniaPas encore d'évaluation
- Field Inspection Techniques Ifad-CaspDocument17 pagesField Inspection Techniques Ifad-CaspNasiru Kura100% (1)
- Latihan MitosisDocument7 pagesLatihan MitosisChe Mahani HussainPas encore d'évaluation
- Microbiological Test of Medical DevicesDocument5 pagesMicrobiological Test of Medical Devicesbijendra_sinhaPas encore d'évaluation
- Biodiversity & StabilityDocument76 pagesBiodiversity & StabilityMAXINE CLAIRE CUTINGPas encore d'évaluation
- Rife S Fundamental Resonance Enhancement A 100 Effective Cure For CancersDocument28 pagesRife S Fundamental Resonance Enhancement A 100 Effective Cure For CancersDimitar LevskiPas encore d'évaluation
- Pain PathwayDocument17 pagesPain PathwaySalsabila Al-BasheerPas encore d'évaluation
- Full Essay Academic SkillsDocument3 pagesFull Essay Academic SkillsNurl AinaPas encore d'évaluation
- Hemostatic Effects of Peperomia PellucidDocument41 pagesHemostatic Effects of Peperomia PellucidGina Rose De GuzmanPas encore d'évaluation
- Transport Phenomena in Nervous System PDFDocument538 pagesTransport Phenomena in Nervous System PDFUdayanidhi RPas encore d'évaluation
- 4 Factors Effecting Shelf Life and Storage Life of Fresh FishDocument26 pages4 Factors Effecting Shelf Life and Storage Life of Fresh FishRafidah Zakaria100% (1)
- Warlock of The Magus World - Arc 2Document808 pagesWarlock of The Magus World - Arc 2Hitsuin MoviesPas encore d'évaluation
- MR Spectroscopy: Technical Aspects and Clinical ApplicationsDocument5 pagesMR Spectroscopy: Technical Aspects and Clinical ApplicationsNotariana Kusuma APas encore d'évaluation
- Evolution of Pharmacy in the PhilippinesDocument2 pagesEvolution of Pharmacy in the PhilippinesRosemarie OngPas encore d'évaluation
- Lipid Movement BiochemDocument9 pagesLipid Movement BiochemCrowPas encore d'évaluation
- 1.4 - Diffusion and Osmosis - CrosswordPuzzleDocument1 page1.4 - Diffusion and Osmosis - CrosswordPuzzlemmssthPas encore d'évaluation
- LearnerDocument7 pagesLearnersudhacarhrPas encore d'évaluation