Vous aimerez peut-être aussi
- Beyond DNA: From Cellular Mechanisms to Environmental Factors: How Epigenetics Shapes Our Biological Destiny and its Implications for Health, Behavior, and the Future of ResearchD'EverandBeyond DNA: From Cellular Mechanisms to Environmental Factors: How Epigenetics Shapes Our Biological Destiny and its Implications for Health, Behavior, and the Future of ResearchPas encore d'évaluation
- Environmental EpigeneticsDocument8 pagesEnvironmental EpigeneticsFrancisco MtzPas encore d'évaluation
- Beyond DNA: The Epigenetic Revolution: From Cellular Mechanisms to Environmental Factors: How Epigenetics Shapes Our Biological Destiny and its Implications for Health, Behavior, and the Future of ResearchD'EverandBeyond DNA: The Epigenetic Revolution: From Cellular Mechanisms to Environmental Factors: How Epigenetics Shapes Our Biological Destiny and its Implications for Health, Behavior, and the Future of ResearchPas encore d'évaluation
- Molecular GeneticDocument15 pagesMolecular Geneticنبأ فؤاد سالمPas encore d'évaluation
- Autism-Spectrum-Disorders-an-Intervention-Approach LAO JIDocument12 pagesAutism-Spectrum-Disorders-an-Intervention-Approach LAO JIErica Alejandra SchumacherPas encore d'évaluation
- Epigenetic Regulation in Exposome-Induced Tumorigenesis-25739Document47 pagesEpigenetic Regulation in Exposome-Induced Tumorigenesis-25739Natalia MinzatuPas encore d'évaluation
- Review Environmental Epigenetics: A Role in Endocrine Disease?Document7 pagesReview Environmental Epigenetics: A Role in Endocrine Disease?Najla MasturaPas encore d'évaluation
- Epigenetics Across The Human LifespanDocument19 pagesEpigenetics Across The Human Lifespanjro84100% (1)
- Nihms 498542Document7 pagesNihms 498542Shrihari WanjalePas encore d'évaluation
- 2013 Proceedings of The Founding Meeting of SF-DOHaDDocument45 pages2013 Proceedings of The Founding Meeting of SF-DOHaDNguyễn Tiến HồngPas encore d'évaluation
- Epigenetics and A New Look On Metabolic Syndrome: ReviewDocument10 pagesEpigenetics and A New Look On Metabolic Syndrome: ReviewrahmaPas encore d'évaluation
- Genes 14 00585 PDFDocument13 pagesGenes 14 00585 PDFandy sotoPas encore d'évaluation
- Written Assign What Is EpigeneticsDocument4 pagesWritten Assign What Is EpigeneticslesleyPas encore d'évaluation
- Epigenetics and Cerebral Organoids Promising DirecDocument12 pagesEpigenetics and Cerebral Organoids Promising DirecViviana ArboledaPas encore d'évaluation
- The Role of Epigenetics in Mental Disorders: Review ArticleDocument7 pagesThe Role of Epigenetics in Mental Disorders: Review ArticleBenet DhasPas encore d'évaluation
- Relationship Between Autism Spectrum Disorder and PesticidesDocument30 pagesRelationship Between Autism Spectrum Disorder and PesticidesJuan Pablo Carrasco SaldiviaPas encore d'évaluation
- Neuro Degenerat yDocument33 pagesNeuro Degenerat yJoel Torres VillenaPas encore d'évaluation
- Environmental Pollution To Blame For Depressive Disorder?: Environmental Research and Public HealthDocument13 pagesEnvironmental Pollution To Blame For Depressive Disorder?: Environmental Research and Public HealthVeroca TorresPas encore d'évaluation
- The Contribution of Environmental Exposure To The Etiology of Autism Spectrum DisorderDocument23 pagesThe Contribution of Environmental Exposure To The Etiology of Autism Spectrum DisorderCristina SolerPas encore d'évaluation
- Novel Di SeasDocument36 pagesNovel Di Seasfarishussein2023Pas encore d'évaluation
- Overview of The Role of Environmental Factors in Neurodevelopmental DisordersDocument18 pagesOverview of The Role of Environmental Factors in Neurodevelopmental DisordersTlaloc GonzalezPas encore d'évaluation
- La Epigenética y Los Estudios en Gemelos en El Campo de La PsiquiatríaDocument9 pagesLa Epigenética y Los Estudios en Gemelos en El Campo de La PsiquiatríaHerb MedrPas encore d'évaluation
- JCM 08 00217 v3Document23 pagesJCM 08 00217 v3Sumit BediPas encore d'évaluation
- Why Schizophrenia Genetics Needs Epigenetics: A ReviewDocument17 pagesWhy Schizophrenia Genetics Needs Epigenetics: A ReviewLinaKoenPas encore d'évaluation
- Epigenética en Trastornos Psiquiátricos 2023Document14 pagesEpigenética en Trastornos Psiquiátricos 2023Sofía Flórez CastilloPas encore d'évaluation
- Jurnal JiwaDocument9 pagesJurnal JiwaRoza EtlaboraPas encore d'évaluation
- Arrowsmith Et Al (2012) - Epigenetic Protein Families A New Frontier For Drug DiscoveryDocument17 pagesArrowsmith Et Al (2012) - Epigenetic Protein Families A New Frontier For Drug DiscoveryRabiatul AdawiyahPas encore d'évaluation
- Sevalia Synthesis PaperDocument20 pagesSevalia Synthesis Paperapi-345271917Pas encore d'évaluation
- Influence of Environmental Exposure On Human Epigenetic RegulationDocument9 pagesInfluence of Environmental Exposure On Human Epigenetic RegulationTiago TorresPas encore d'évaluation
- 2006-SKELTON Obesity and Its Therapy From Genes To Community ActionDocument17 pages2006-SKELTON Obesity and Its Therapy From Genes To Community ActionNovi LatifaPas encore d'évaluation
- AdvancedDocument6 pagesAdvancedMustafa HatemPas encore d'évaluation
- Understanding Insulin in The Age of Precision Medicine and Big Data - Under-Explored Nature of GenomicsDocument38 pagesUnderstanding Insulin in The Age of Precision Medicine and Big Data - Under-Explored Nature of GenomicsGestne AurePas encore d'évaluation
- Nutri (Epi) Genomics and Metabolic SyndromeDocument12 pagesNutri (Epi) Genomics and Metabolic SyndromeLuama AraújoPas encore d'évaluation
- Juruena 2021Document9 pagesJuruena 2021PailonCPas encore d'évaluation
- Endocrine Disrupting Chemicals and Disease SusceptibilityDocument28 pagesEndocrine Disrupting Chemicals and Disease SusceptibilityTiago TorresPas encore d'évaluation
- Flores Dorantes 27 Environment and Gene Association With Obesity and Their Impact On Neurodegenerative and Neurodevelopmental DiseasesDocument24 pagesFlores Dorantes 27 Environment and Gene Association With Obesity and Their Impact On Neurodegenerative and Neurodevelopmental DiseasesPaty BritoPas encore d'évaluation
- Gene Environment InteractionDocument26 pagesGene Environment InteractionAwais GulPas encore d'évaluation
- tmpED54 TMPDocument6 pagestmpED54 TMPFrontiersPas encore d'évaluation
- Autistic Spectrum Disorders As Functional Disconnection SyndromeDocument53 pagesAutistic Spectrum Disorders As Functional Disconnection SyndromeFrontiersPas encore d'évaluation
- Evolutionary Theory Essay - 74Document7 pagesEvolutionary Theory Essay - 74frazzycakebossPas encore d'évaluation
- FunctionalDisconnectionSyndrome MelilloLeismanMay09Document53 pagesFunctionalDisconnectionSyndrome MelilloLeismanMay09Brain Balance Georgia100% (1)
- Farmakogenetik AdoDocument31 pagesFarmakogenetik AdoEndang SusilowatiPas encore d'évaluation
- Oral Inflammation, Tooth Loss, Risk Factors, and Association With Progression of Alzheimer's DiseaseDocument44 pagesOral Inflammation, Tooth Loss, Risk Factors, and Association With Progression of Alzheimer's DiseaseAndrea Cabibbo Fisioterapista-OsteopataPas encore d'évaluation
- En 24 257Document16 pagesEn 24 257Trinni GuzmanPas encore d'évaluation
- Review Article: Pharmacogenomics of Drug Response in Type 2 Diabetes: Toward The Definition of Tailored Therapies?Document11 pagesReview Article: Pharmacogenomics of Drug Response in Type 2 Diabetes: Toward The Definition of Tailored Therapies?James CowellPas encore d'évaluation
- Liu2020 Article GutMicrobiotaAndDysbiosisInAlzDocument18 pagesLiu2020 Article GutMicrobiotaAndDysbiosisInAlzMonaPas encore d'évaluation
- Immune System An Emerging Player of EDC Effect On Metabolic HealthDocument14 pagesImmune System An Emerging Player of EDC Effect On Metabolic Healthshweta pasiPas encore d'évaluation
- Food & Genes: Term Paper By: Phurpa D. ThungonDocument15 pagesFood & Genes: Term Paper By: Phurpa D. ThungonMehul BhardwajPas encore d'évaluation
- Accepted Manuscript: Reproductive ToxicologyDocument45 pagesAccepted Manuscript: Reproductive ToxicologyAline SquassoniPas encore d'évaluation
- Epigenetics: A Possible Answer To The Undeciphered Etiopathogenesis and Behavior of Oral LesionsDocument8 pagesEpigenetics: A Possible Answer To The Undeciphered Etiopathogenesis and Behavior of Oral LesionsEstaf EmkeyzPas encore d'évaluation
- Endocrine Disrupters - A Review of Some Sources, Effects, and Mechanisms of Actions On Behaviour and Neuroendocrine SystemsDocument28 pagesEndocrine Disrupters - A Review of Some Sources, Effects, and Mechanisms of Actions On Behaviour and Neuroendocrine SystemsTiago TorresPas encore d'évaluation
- 404 Assignmet Roll SH 08Document6 pages404 Assignmet Roll SH 08Mobashir RahmanPas encore d'évaluation
- SampleafdfsfsdgfgdfgsDocument1 pageSampleafdfsfsdgfgdfgsLinba YtPas encore d'évaluation
- Epigenetics and Aging: ReviewDocument20 pagesEpigenetics and Aging: ReviewDeepak PanwarPas encore d'évaluation
- Nutrients: Nutrigenetics and Nutrigenomics Insights Into Diabetes EtiopathogenesisDocument32 pagesNutrients: Nutrigenetics and Nutrigenomics Insights Into Diabetes EtiopathogenesisPuji HananiPas encore d'évaluation
- The Ketogenic Diet As A Non Pharmacological Treatment For HIV Associated Neurocognitive Disorder A Descriptive AnalysisDocument5 pagesThe Ketogenic Diet As A Non Pharmacological Treatment For HIV Associated Neurocognitive Disorder A Descriptive AnalysisPaul HartingPas encore d'évaluation
- Rationale For Dietary Antioxidant Treatment of ADHDocument20 pagesRationale For Dietary Antioxidant Treatment of ADHFernando SouzaPas encore d'évaluation
- Biomolecules 11 00543Document19 pagesBiomolecules 11 00543gamers juarezPas encore d'évaluation
- Toxicology: M.F. Simoniello, L. Contini, E. Benavente, C. Mastandrea, S. Roverano, S. PairaDocument7 pagesToxicology: M.F. Simoniello, L. Contini, E. Benavente, C. Mastandrea, S. Roverano, S. PairaMauro Porcel de PeraltaPas encore d'évaluation
- Jurnal AutismDocument10 pagesJurnal AutismAby SuryaPas encore d'évaluation
- Disruptive Mood Dysregulation Disorder in ICD 11: A New Disorder or ODD With A Specifier For Chronic Irritability?Document2 pagesDisruptive Mood Dysregulation Disorder in ICD 11: A New Disorder or ODD With A Specifier For Chronic Irritability?Rafael MartinsPas encore d'évaluation
- Research in Developmental DisabilitiesDocument9 pagesResearch in Developmental DisabilitiesRafael MartinsPas encore d'évaluation
- Wax Mon Sky 2017Document22 pagesWax Mon Sky 2017Rafael MartinsPas encore d'évaluation
- The History, Diagnosis and Treatment of Disruptive Mood Dysregulation DisorderDocument4 pagesThe History, Diagnosis and Treatment of Disruptive Mood Dysregulation DisorderRafael MartinsPas encore d'évaluation
- Ogasawara 2017Document12 pagesOgasawara 2017Rafael MartinsPas encore d'évaluation
- HHS Public Access: Cognitive-Behavioral Therapy For A 9-Year-Old Girl With Disruptive Mood Dysregulation DisorderDocument20 pagesHHS Public Access: Cognitive-Behavioral Therapy For A 9-Year-Old Girl With Disruptive Mood Dysregulation DisorderRafael MartinsPas encore d'évaluation
- NIH Public Access: Childhood Maltreatment, Emotional Dysregulation, and Psychiatric ComorbiditiesDocument20 pagesNIH Public Access: Childhood Maltreatment, Emotional Dysregulation, and Psychiatric ComorbiditiesRafael MartinsPas encore d'évaluation
- Ni Hms 671626Document9 pagesNi Hms 671626Rafael MartinsPas encore d'évaluation
- HHS Public Access: DSM-5 Disruptive Mood Dysregulation Disorder: Correlates and Predictors in Young ChildrenDocument21 pagesHHS Public Access: DSM-5 Disruptive Mood Dysregulation Disorder: Correlates and Predictors in Young ChildrenRafael MartinsPas encore d'évaluation
- NIH Public Access: DSM-5: Disruptive Mood Dysregulation DisorderDocument11 pagesNIH Public Access: DSM-5: Disruptive Mood Dysregulation DisorderRafael MartinsPas encore d'évaluation
- Ni Hms 645602Document21 pagesNi Hms 645602Rafael MartinsPas encore d'évaluation
- HHS Public Access: Emotion Dysregulation and Negative Affect: Association With Psychiatric SymptomsDocument18 pagesHHS Public Access: Emotion Dysregulation and Negative Affect: Association With Psychiatric SymptomsRafael MartinsPas encore d'évaluation
- HHS Public Access: Irritability in Children and Adolescents: Past Concepts, Current Debates, and Future OpportunitiesDocument13 pagesHHS Public Access: Irritability in Children and Adolescents: Past Concepts, Current Debates, and Future OpportunitiesRafael MartinsPas encore d'évaluation
- NIH Public Access: Author ManuscriptDocument19 pagesNIH Public Access: Author ManuscriptRafael MartinsPas encore d'évaluation
- NIH Public Access: Adult Diagnostic and Functional Outcomes of DSM-5 Disruptive Mood Dysregulation DisorderDocument15 pagesNIH Public Access: Adult Diagnostic and Functional Outcomes of DSM-5 Disruptive Mood Dysregulation DisorderRafael MartinsPas encore d'évaluation
- Summary Notes - Topic 9 Transport in Animals - CIE Biology IGCSEDocument5 pagesSummary Notes - Topic 9 Transport in Animals - CIE Biology IGCSEMadhan Reddy SuddalaPas encore d'évaluation
- My Own Life. Oliver SacksDocument3 pagesMy Own Life. Oliver Sacksajjm100% (4)
- Auras PDFDocument24 pagesAuras PDFvijayant pandeyPas encore d'évaluation
- PletizmografijaDocument29 pagesPletizmografijaJelenaJankovPas encore d'évaluation
- Community Health Nursing POST TEST Day 1Document1 pageCommunity Health Nursing POST TEST Day 1Mcbry TiongPas encore d'évaluation
- Pyelonephritis: Dr. Shraddha Koirala Department of Pathology NMCTHDocument26 pagesPyelonephritis: Dr. Shraddha Koirala Department of Pathology NMCTHUrusha MagarPas encore d'évaluation
- Calamansi and Lemongrass As Alternative Mosquito Coil: Submitted By: Kristhel Nieves N. Zamora 10-AmethystDocument6 pagesCalamansi and Lemongrass As Alternative Mosquito Coil: Submitted By: Kristhel Nieves N. Zamora 10-AmethystKristhel ZamoraPas encore d'évaluation
- Santosh DevDocument45 pagesSantosh Devdevdsantosh100% (1)
- Glossary of Commonly Used Dental Terms: 14212 Ambaum Boulevard Southwest Suite #101, Burien, WA 98166Document10 pagesGlossary of Commonly Used Dental Terms: 14212 Ambaum Boulevard Southwest Suite #101, Burien, WA 98166Mayra RojasPas encore d'évaluation
- The Importance of SleepDocument4 pagesThe Importance of Sleep7A323Trọng NhânPas encore d'évaluation
- Vestibular SchwannomaDocument12 pagesVestibular SchwannomaEustakia Rini Kartika DewiPas encore d'évaluation
- Pharmacology of Drugs Acting On Renal SystemDocument20 pagesPharmacology of Drugs Acting On Renal SystemBestha Chakri100% (2)
- SaveBooklet BookletDocument120 pagesSaveBooklet BookletRoscelie KhoPas encore d'évaluation
- Surgery Ospe 322Document322 pagesSurgery Ospe 322fbf50269Pas encore d'évaluation
- Saudi License Exam (SLE) For General Dentistry - Sep 6, 2011Document10 pagesSaudi License Exam (SLE) For General Dentistry - Sep 6, 2011asifhameed90% (10)
- Unidirctional Anti Covid-19 Gate Using Motion SensorDocument45 pagesUnidirctional Anti Covid-19 Gate Using Motion SensorMuhammad SalmanPas encore d'évaluation
- دليل كامل عن صناعة جبنة الموزريلاDocument56 pagesدليل كامل عن صناعة جبنة الموزريلاyahyaalmoalmPas encore d'évaluation
- Street Corner Secrets by Svati ShahDocument71 pagesStreet Corner Secrets by Svati ShahDuke University Press100% (2)
- 2020 11 07 XII Economics 1Document17 pages2020 11 07 XII Economics 1Sjft6hd FfPas encore d'évaluation
- Botany ProjectDocument25 pagesBotany ProjectSunil ShekharPas encore d'évaluation
- Neonatal HyperglycemiaDocument18 pagesNeonatal HyperglycemiayumicheongPas encore d'évaluation
- Chromosome Walking: and David S. Hogness in The Early 1980'sDocument4 pagesChromosome Walking: and David S. Hogness in The Early 1980'sprakash singhPas encore d'évaluation
- 299.08 - Renal Pathology) Chronic Kideny Disease (CKD)Document15 pages299.08 - Renal Pathology) Chronic Kideny Disease (CKD)AnishilPas encore d'évaluation
- Transgenic AnimaliaDocument4 pagesTransgenic AnimaliaGalino Julia Cristine A.Pas encore d'évaluation
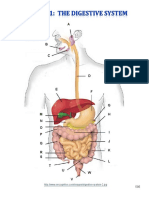
- Digestive System Charts 1Document14 pagesDigestive System Charts 1api-306762527Pas encore d'évaluation
- American Family PhysicianDocument4 pagesAmerican Family PhysicianGoddess Nth The Artemis EffectPas encore d'évaluation
- TRS601 Vocabulary ListDocument5 pagesTRS601 Vocabulary ListNgo Van Hung (K17 HCM)Pas encore d'évaluation
- Mental Health DirectoryDocument88 pagesMental Health DirectoryAbhijit DasPas encore d'évaluation
- Active Fluid De-Resuscitacion in Crtiticalli III Patients Whitj ShockDocument8 pagesActive Fluid De-Resuscitacion in Crtiticalli III Patients Whitj ShockMartha Isabel BurgosPas encore d'évaluation
- Symptoms and Signs: Compartment SyndromeDocument4 pagesSymptoms and Signs: Compartment SyndromeajeikoPas encore d'évaluation
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesD'EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesÉvaluation : 4.5 sur 5 étoiles4.5/5 (397)
- Gut: the new and revised Sunday Times bestsellerD'EverandGut: the new and revised Sunday Times bestsellerÉvaluation : 4 sur 5 étoiles4/5 (393)
- Why We Die: The New Science of Aging and the Quest for ImmortalityD'EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityÉvaluation : 4 sur 5 étoiles4/5 (5)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceD'EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceÉvaluation : 4.5 sur 5 étoiles4.5/5 (517)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisD'EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisÉvaluation : 3.5 sur 5 étoiles3.5/5 (2)
- Masterminds: Genius, DNA, and the Quest to Rewrite LifeD'EverandMasterminds: Genius, DNA, and the Quest to Rewrite LifePas encore d'évaluation
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessD'Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessÉvaluation : 4 sur 5 étoiles4/5 (33)
- Tales from Both Sides of the Brain: A Life in NeuroscienceD'EverandTales from Both Sides of the Brain: A Life in NeuroscienceÉvaluation : 3 sur 5 étoiles3/5 (18)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsD'EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsÉvaluation : 4.5 sur 5 étoiles4.5/5 (6)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionD'EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionÉvaluation : 4 sur 5 étoiles4/5 (812)
- Return of the God Hypothesis: Three Scientific Discoveries That Reveal the Mind Behind the UniverseD'EverandReturn of the God Hypothesis: Three Scientific Discoveries That Reveal the Mind Behind the UniverseÉvaluation : 4.5 sur 5 étoiles4.5/5 (52)
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperD'EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperÉvaluation : 4.5 sur 5 étoiles4.5/5 (16)
- Who's in Charge?: Free Will and the Science of the BrainD'EverandWho's in Charge?: Free Will and the Science of the BrainÉvaluation : 4 sur 5 étoiles4/5 (65)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)D'EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Évaluation : 4 sur 5 étoiles4/5 (411)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedD'EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedÉvaluation : 4 sur 5 étoiles4/5 (11)
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomD'EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomÉvaluation : 4 sur 5 étoiles4/5 (216)
- Good Without God: What a Billion Nonreligious People Do BelieveD'EverandGood Without God: What a Billion Nonreligious People Do BelieveÉvaluation : 4 sur 5 étoiles4/5 (66)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorD'EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorPas encore d'évaluation
- Seven and a Half Lessons About the BrainD'EverandSeven and a Half Lessons About the BrainÉvaluation : 4 sur 5 étoiles4/5 (110)
- Human: The Science Behind What Makes Your Brain UniqueD'EverandHuman: The Science Behind What Makes Your Brain UniqueÉvaluation : 3.5 sur 5 étoiles3.5/5 (38)
- Moral Tribes: Emotion, Reason, and the Gap Between Us and ThemD'EverandMoral Tribes: Emotion, Reason, and the Gap Between Us and ThemÉvaluation : 4.5 sur 5 étoiles4.5/5 (115)
- The Rise and Fall of the Dinosaurs: A New History of a Lost WorldD'EverandThe Rise and Fall of the Dinosaurs: A New History of a Lost WorldÉvaluation : 4 sur 5 étoiles4/5 (595)
- Minds Make Societies: How Cognition Explains the World Humans CreateD'EverandMinds Make Societies: How Cognition Explains the World Humans CreateÉvaluation : 4.5 sur 5 étoiles4.5/5 (24)
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouD'EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouÉvaluation : 4.5 sur 5 étoiles4.5/5 (62)
- The Invention of Tomorrow: A Natural History of ForesightD'EverandThe Invention of Tomorrow: A Natural History of ForesightÉvaluation : 4.5 sur 5 étoiles4.5/5 (5)
- The Lives of Bees: The Untold Story of the Honey Bee in the WildD'EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildÉvaluation : 4.5 sur 5 étoiles4.5/5 (44)