Académique Documents
Professionnel Documents
Culture Documents
U2
Transféré par
Matías BacherTitre original
Copyright
Formats disponibles
Partager ce document
Partager ou intégrer le document
Avez-vous trouvé ce document utile ?
Ce contenu est-il inapproprié ?
Signaler ce documentDroits d'auteur :
Formats disponibles
U2
Transféré par
Matías BacherDroits d'auteur :
Formats disponibles
Instituto Hernando Arias de Saavedra
BIOLOGA
1
A AP PU UN NT TE ES S D DE E F FU UN ND DA AM ME EN NT TO OS S D DE E B BI IO OL LO OG G A A
P Pr ro of fe es so or ra ad do o e en n B Bi io ol lo og g a a
T Te ec cn ni ic ca at tu ur ra a e en n G Ge es st ti i n n A Am mb bi ie en nt ta al l
U Un ni id da ad d 2 2
Instituto Hernando Arias de Saavedra
BIOLOGA
2
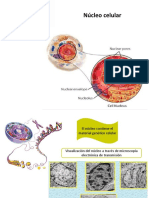
E L N C L E O
El ncleo es un compartimento intracelular tpico de las clulas eucariotas.
Contiene al ADN. Est formado por la envuelta nuclear y por el nucleoplasma.
En la envuelta nuclear se encuentran los poros nucleares, puertas de entrada y de
salida del ncleo.
El ADN se dispone en el nucleoplasma formando la cromatina: ADN ms protenas
asociadas
El ncleo es la estructura que caracteriza a las clulas eucariotas. Es el compartimento
donde se encuentra el ADN y toda la maquinaria necesaria para transcribir su informacin
a ARN y para replicarse.
Normalmente aparece un solo ncleo por clula, aunque en algunos casos hay ms de
uno como ocurre en los osteoclastos, en las fibras musculares esquelticas o en los
epitelios de algunos invertebrados. La forma nuclear es variable y se suele adaptar a la
forma celular, aunque no siempre es as. Por ejemplo, los neutrfilos de la sangre poseen
ncleos multilobulados. La localizacin habitual del ncleo es en el centro de la clula,
pero tambin puede situarse en otras posiciones ms perifricas. As, en las clulas
secretoras se puede localizar en la parte basal de la clula y en las musculares
esquelticas se dispone en las proximidades de la membrana plasmtica.
Distintos tipos de ncleos. A. Clulas epiteliales de la vescula biliar de humanos con los ncleos
redondeados. B Monocito de la sangre con el ncleo arrionado. C. Neutrfilos de la sangre con los ncleos
multilobulados. D. Vista parcial de una clula muscular multinucleada, con los ncleos situados en zona
perifrica (flechas).
Instituto Hernando Arias de Saavedra
BIOLOGA
3
El ncleo consta de dos componentes que se pueden distinguir morfolgicamente: la
envuelta o membrana nuclear y el nucleoplasma.
La envuelta o membrana nuclear separa el nucleoplasma del citoplasma. En ella
se encuentran las puertas que comunican estos dos espacios, los poros
nucleares, que permiten el intercambio de molculas en los dos sentidos pero de
una manera especfica y regulada.
En el nucleoplasma se encuentra:
el ADN y sus protenas asociadas formando la
cromatina, que si est muy compactada se le
denomina heterocromatina y si aparece ms laxa
se le denomina eucromatina.
y el nuclolo.
En este eje vamos a tratar todos los procesos relacionados con la transcripcin y la
regulacin gnica, as como una parte relacionada con la organizacin del ADN y de los
genes que lo forman. Trataremos la morfologa nuclear y te aconsejamos buscar
informacin sobre el ADN en textos o sitios web relacionados con la gentica.
L A E N V U E L T A N U C L E A R
La envuelta o membrana nuclear separa el nucleoplasma del citoplasma.
Est formada por una membrana interna, una membrana externa, un espacio
intermembrana, por los poros nucleares y por la lmina nuclear.
Es responsable de la estructura y de la forma del ncleo, gracias sobre todo a la lmina
nuclear.
Regula el trasiego de molculas entre el nucleoplasma y el citoplasma, que est mediado
por los poros nucleares.
La membrana externa se contina con la membrana del retculo endoplasmtico
La envuelta nuclear separa fsicamente al nucleoplasma (cromatina y dems
componentes del interior nuclear) del citoplasma y regula el movimiento de
macromolculas entre ellos, establece la forma nuclear y contribuye a la organizacin
interna del ncleo ya que aporta lugares de anclaje para la cromatina.
Instituto Hernando Arias de Saavedra
BIOLOGA
4
La envuelta nuclear est formada por una membrana doble, externa e interna, quedando
entre ambas un espacio intermembranoso (de aproximadamente 25-40 nm), formando
todos estos elementos las denominas cisternas perinucleares.
La membrana externa se contina con la del retculo endoplasmtico y posee ribosomas
adheridos. La membrana interna contiene una composicin molecular diferente y posee
protenas transmembrana que interactan con la cromatina y con la lmina nuclear, el otro
componente de la envuelta nuclear. Existe una comunicacin entre la membrana nuclear
interna y externa en la periferia de los poros nucleares.
Qu mantiene las diferencias en la composicin de ambas membranas?
Parece existir un mecanismo de retencin selectiva. Las protenas se sintetizan en el
retculo endoplasmtico que llegan a la membrana interna por difusin, pero slo aquellas
que interaccionan con las protenas de la lmina nuclear o de la cromatina se mantienen
aqu.
Esquema de la estructura de la envuelta nuclear. Est formada por una membrana externa, por el espacio
intermembrana, por la membrana interna y por la lmina nuclear. La membrana externa se contina con el
retculo endoplasmtico. Los poros nucleares se encuentran insertos en interrupciones puntuales de la
envuelta nuclear.
La lmina o membrana nuclear es un entramado proteico que separa la membrana
interna de la envuelta nuclear de la cromatina, (en mamferos tiene un espesor de 20 a 25
nm). Las principales protenas que la componen se denominan lminas, que se
encuentran en dos isoformas: tipo A (lminas A y C) y tipo B (lminas B1 y B2/3).
Instituto Hernando Arias de Saavedra
BIOLOGA
5
Pertenecen a la familia de los filamentos intermedios. Estas protenas se disponen en
forma de malla cubriendo toda la cara interna de la envuelta nuclear a la cual estn
unidas por un lado y por el otro a la cromatina. La asociacin ntima entre la membrana
interna de la envuelta nuclear y la lmina nuclear se produce gracias a la existencia de al
menos 20 protenas localizadas en la membrana interna.
Una de sus principales funciones de la lmina nuclear es la de mantener la estructura de
la envuelta nuclear. Las deficiencias en estas protenas producen las enfermedades
denominadas laminopatas
1
, las cuales presentan ncleos desorganizados y pueden
llevar a la muerte celular o a la fragilidad de la envuelta nuclear. La desorganizacin y
organizacin de la envuelta nuclear durante la divisin celular se debe a acciones
enzimticas sobre las lminas. La forma nuclear cambia cuando cambia la expresin de
las protenas que forman la lmina nuclear, lo cual es observable durante el desarrollo
embrionario, la diferenciacin celular o ciertas patologas celulares. Otra de las funciones
de la lmina nuclear es servir de soporte para diversas reacciones relacionadas con la
cromatina. Pero, adems sirve de punto de anclaje del ncleo al citoesqueleto de la
clula, lo que permite al ncleo situarse en una posicin determinada dentro de la clula o
moverse por su interior.
En la envuelta nuclear se encuentran los poros nucleares, que son los responsables de
controlar el trasiego de molculas entre el interior del ncleo y el citoplasma, y que
veremos en el siguiente apartado.
Por qu una clula necesita separar el ADN del citoplasma, cuando esto le
supone un considerable consumo de recursos?
Entre las razones ms evidentes estn:
a) Estabilidad gnica: la confinacin del genoma en un compartimento contribuye a
preservar la estabilidad del ADN, que es mayor que en procariotas, teniendo en cuenta
que estamos hablando de una enorme cantidad de ADN.
1
Las laminopatas son un conjunto de enfermedades raras que comparten formas errneas de codificacin
gentica de las lminas, protenas constitutivas de la lmina nuclear. Son trastornos que afectan a diferentes
tejidos y funciones como el tejido muscular estriado, el tejido adiposo, el tejido seo, el sistema nervioso o el
envejecimiento precoz. La distrofia muscular de Emery-Dreifuss o el sndrome Hutchinson-Gildford Progeria
son ejemplos de laminopatas. Conjuntamente con ellas suelen estudiarse otros trastornos que provocan de
forma directa o indirecta alteracin de la funcin de las lminas. El creciente inters y estudio que las
laminopatas han suscitado desde hace una dcada podran permitir encontrar respuestas a preguntas clave
en relacin con el metabolismo, la resistencia a la insulina, la muerte sbita o el envejecimiento. En este
trabajo se revisan las laminopatas desde un punto de vista molecular, patognico y clnico.
Instituto Hernando Arias de Saavedra
BIOLOGA
6
b) Permite la regulacin de la expresin gnica a un nivel impensable para procariotas.
Por ejemplo, el acceso o no a factores de transcripcin. Los factores de transcripcin son
molculas que regulan la expresin gnica y son sintetizados en el citoplasma. Para su
accin deben ser transportados al interior celular. Las cascadas de sealizacin empiezan
en receptores de membrana o internos, pero cualquiera que sea su inicio, si
desencadenan expresin gnica, alguna molcula de la cascada de sealizacin debe
atravesar la envuelta nuclear. Si se bloquea este paso no se producir ningn efecto
sobre la expresin gnica.
c) La presencia de intrones y exones en los genes eucariotas obliga a una maduracin del
transcrito primario. Es muy peligroso que un ARNm sin madurar acceda a los ribosomas
puesto que producira protenas no funcionales o incluso potencialmente peligrosas,
d) Separar la transcripcin de la traduccin aporta a la clula una herramienta ms para
regular la informacin que va desde el ADN hasta la protena. As, la transcripcin de un
gen a ARNm no significa que se produzca una protena de forma inmediata. Impidiendo la
salida del ARNm del ncleo se evita la produccin de dicha protena.
Bibliografa especfica: Rowat AC, Lammerding J, Herrmann H, Aebi U . Towards and
integrated understanding of the structure and mechanics of the cell nucleus. 2008.
BioEssays 30: 226-236.
P O R O S N U C L E A R E S
Los poros nucleares son complejos proteicos localizados en la envuelta nuclear cuya
misin es permitir y controlar el trfico de molculas entre el nucleoplasma y el
citoplasma.
Las protenas que forman los poros nucleares se denominan nucleoporinas.
El transporte mediado por los poros nucleares es especfico.
Las importinas y las exportinas son dos de las familias de molculas que reconocen
pptidos seal de entrada o salida, respectivamente, de aquellas molculas que tienen
que entrar o salir del ncleo. Son estas dos familias de protenas las que interaccionan
con las nucleoporinas durante su trasiego por los poros nucleares.
Instituto Hernando Arias de Saavedra
BIOLOGA
7
Las cisternas de la envuelta nuclear (dos membranas ms el especio entre ambas), dejan
unos huecos entre ellas donde se encuentran los poros nucleares. stos son grandes
complejos moleculares visibles con el microscopio electrnico, y denominados en su
conjunto complejo del poro. Son la puerta de comunicacin entre ncleo y citoplasma y
todo el transporte nucleocitoplasmtico se da a travs de ellos.
Hay que tener en cuenta que no son estructuras transmembrana como ocurra con las
protenas integrales, sino que se podran considerar como complejos proteicos asociados
a las membranas de la envuelta nuclear. Los poros nucleares son muy numerosos en las
clulas que requieren un alto trnsito de sustancias entre el ncleo y el citoplasma como
por ejemplo en las clulas que se estn diferenciando. Se considera que puede haber
unos 11 poros por m2, lo que equivale a unos 3000 a 4000 poros por ncleo. El dimetro
de un poro nuclear es de unos 80 nm pero el espacio til para el trasiego de las molculas
que se transportan es de
unos 50 nm.
Esquema de la estructura proteica
de los poros nucleares.
(Modificado de Beck et al., 2007)
Las protenas que forman parte del complejo del poro se denominan nucleoporinas. En
las levaduras hay unas 30 protenas distintas en cada poro nuclear, mientras que en los
metazoos pueden ser 40 o ms. Se cree que un poro de una clula de mamferos
contiene unas 400 nucleoporinas, ya que muchas de ellas se encuentran repetidas. Las
protenas que forman los poros nucleares se asocian para formar 8 bloques que
configuran un octgono regular y se organizan formando anillos: el anillo citoplasmtico
orientado hacia el citoplasma, el anillo radial situado en el hueco que deja la envuelta
nuclear y es responsable de anclar el complejo del poro a las membranas de la envuelta
nuclear y el anillo nuclear que se encuentra en el nucleoplasma.
Adems, desde cada bloque se proyectan fibrillas proteicas que van hacia el citoplasma
denominadas fibras citoplasmticas, y otras al interior del ncleo que reciben el nombre
Instituto Hernando Arias de Saavedra
BIOLOGA
8
de fibras nucleares. stas ltimas se conectan a otro conjunto de protenas que forman
una estructura cerrada llamada jaula nuclear o anillo distal.
Los poros nucleares contienen un canal que se puede expandir cuando realiza transporte
activo. El canal puede permitir el paso libre de pequeas molculas pero restringir el
movimiento de otras de mayor tamao. Aunque incluso algunas molculas menores tales
como las histonas, los ARNt o algunos ARNm. Estas molculas y otras ms grandes
cruzan por transporte pasivo facilitado. Pero aunque sea pasivo facilitado pueden viajar
en contra de su gradiente, por tanto necesitan energa que es aportada por el gradiente
de otras molculas denominadas Ran-GTPasas. Veamos como ocurre.
Gradientes creados por las molculas Ran entre el citoplasma y el nucleoplasma. La energa se consume en
el nucleoplasma para crear Ran-GTP a partir de Ran y fsforo inorgnico, manteniendo la concentracin de
Ran-GTP elevada. En el citoplasma las Ran-GTP son rpidamente convertidas en Ran-GDP, manteniendo la
concentracin de estas ltimas elevada. Mientras, la hidrlisis del Ran-GDP a Ran, mantiene la concentracin
de Ran-GDP baja en el nucleoplasma. El tamao de los iconos son indicativos de la concentracin.
El transporte mediado por los poros nucleares est orquestado por las protenas Ran-
GTPasas y por unas familias de protenas denominadas importinas y exportinas. Las
molculas Ran-GTPasas son trascendentales tanto para la importacin como para la
exportacin de molculas. Son las responsables de crear un gradiente que dirige el
transporte y crear este gradiente es la nica parte del transporte por parte de los poros
nucleares que gasta energa. Las molculas Ran pueden estar en tres estados: Ran-GTP,
Ran-GDP y Ran. El paso de un estado a otro est mediado por otras enzimas. En el
nucleoplasma hay una mayor concentracin de Ran-GTP, mientras que en el citoplasma
abunda la Ran-GTP.
Instituto Hernando Arias de Saavedra
BIOLOGA
9
Las protenas que tienen que ser importadas al nucleoplasma necesitan poseer una
secuencia de aminocidos o pptido seal de entrada y las que tienen que ser
exportadas, un pptido da la seal de salida. Estas secuencias de aminocidos sern
reconocidas por las importinas o por las exportinas, respectivamente. Las protenas de los
poros nucleares no interaccionan directamente con las protenas transportadas sino son
las importinas y las exportinas.
Estos complejos importina-protena o exportina-protena utilizan los gradientes de las
molculas Ran-GTP o Ran-GDP para cruzar los poros nucleares y llevar sus cargas al
otro lado. Adems de protenas, las molculas de ARN deben tambin atravesar los poros
nucleares. El mecanismo que usan los distintos tipos de ARN para ser transportados
difieren entre s, pero todos estn mediados por un mecanismo de asociacin con
protenas. El exporte de ARNt sigue un mecanismo en el que es reconocido por una
exportina denominada exportina-t, que tambin se une a una Ran-GTP. El mecanismo de
exportacin de los ARNr no se conoce muy bien. Los ARNm no utiliza el sistema de Ran-
GTP sino que su transporte est mediado por dos protenas que forman el complejo
Tap/Nxt, el cual interacciona con las nucleoporinas.
Bibliografa especfica: Beck M, Lucic V, Forster F, Baumeister W, Medalia O .
Snapshots of nuclear pore complexes in action captured by cryo-electron tomography.
2007. Nature 449:611-615.
COMPLEJOS DE PORO NUCLEAR
La envoltura nuclear presenta estructuras discoidales llamadas complejos de poro
nuclear (CPN)
El nmero de CPN es variable, incrementndose a medida que aumenta la actividad
celular. En una clula de mamfero hay entre 3000 a 4000 complejos de poro. Cada CPN
es una estructura macromolecular compleja constituida por un gran nmero de protenas
de disposicin octamrica. Est formado por:
Ocho columnas proteicas, que forman las paredes laterales del poro.
Un anillo externo, formado por ocho unidades proteicas.
Un anillo interno, tambin con estructura octamrica.
Protenas de anclaje que fijan cada columna al espacio perinuclear.
Protenas radiales que se proyectan desde las columnas hacia la luz del poro, a
manera de diafragma
Instituto Hernando Arias de Saavedra
BIOLOGA
10
Protenas fibrilares fijas al anillo interno y externo. En la cara nuclear convergen
para formar una canastilla o cesta. A lo largo de estas fibrillas se ubican
nucleoporinas que intervienen en el transporte de sustancias a travs del poro.
Un poro central o abertura.
Esquema del complejo de poro nuclear
La luz de los CPN suele presentarse obturada por las protenas que circulan a travs del
poro, como por las carioportinas (Kap) que actan como eficientes transportadores en el
trfico ncleo/citoplasma.
Los CPN presentan uno o varios canales acuosos a travs de los cuales las pequeas
molculas solubles en agua difunden (transporte no regulado). Las molculas de mayor
peso molecular son transportadas en forma activa, por lo que requieren energa y
molculas transportadoras.
Se importan dentro del ncleo:
Las protenas sintetizadas en el citoplasma necesarias para ensamblar los
ribosomas.
Los factores de transcripcin requeridos en la activacin o inactivacin de los
genes.
Los factores de empalme necesarios en el proceso de maduracin de los
ribosomas.
Instituto Hernando Arias de Saavedra
BIOLOGA
11
Las molculas y macromolculas ensambladas y exportadas desde el ncleo al
citoplasma incluyen:
Las subunidades ribosomales
ARNm
ARN de transferencia
Factores de transcripcin que son devueltos al citoplasma para ser reutilizados.
Los mecanismos implicados en el transporte a travs del poro son diferentes al transporte
de protenas en las membranas de otras organelas. Por ejemplo, las protenas nucleares
son transportadas a travs del poro manteniendo su conformacin plegada, por el
contrario las protenas que no se localizarn en el ncleo se despliegan durante el
transporte.
Los complejos de poro nuclear hacen de la envoltura nuclear una barrera selectiva entre
el ncleo y el citoplasma. Estos complejos constituyen la principal va de comunicacin
entre el compartimiento nuclear y citoplasmtico de la clula ante el pesado trfico
molecular. Aun cuando las protenas pequeas y otras molculas viajan a travs de los
canales perifricos, las protenas de gran tamao deben poseer una etiqueta para
ingresar por el canal central. Estas protenas sintetizadas en el citoplasma contienen la
seal de localizacin nuclear (nuclear signal localization, NSL).
Tampoco los ARN pueden salir de ncleo por s mismos. Ellos salen a travs del
complejo de poro con una protena especial que posee una seal nuclear de exportacin
(nuclear export signal, NES). Ambas NSL y NES consisten en una secuencia corta de
aminocidos, muchos de los cuales tienden a estar ubicados centralmente dentro de las
protenas.
Las proyecciones filamentosas desde la cara citoslica del complejo pueden enlazar
protenas, conducindolas dentro del poro central. Los filamentos citoslicos, el poro
central y la canasta nuclear se proyectan dentro del compartimiento nuclear. Se cree que
la canasta puede ser un rea importante de paso para la preparacin de las
ribonucleoprotenas (RNP) antes de ser exportadas.
Las molculas de mayor tamao requieren de una protena transbordadora o
carioportina. La superfamilia de carioportinas est integrada por: importinas . exportinas
y transportinas .
Instituto Hernando Arias de Saavedra
BIOLOGA
12
EXPORTACIN DE ARN
Los ARN maduros se asocian a protenas llamadas transportinas, las cuales actan como
transbordadores permitiendo el pasaje de ARN al citoplasma. En la siguiente figura se
esquematiza como el ARNm es llevado fuera del ncleo. Los ARNm maduros que
presentan la poli A se asocian con varias protenas, formando una partcula de
ribonucleoprotena (RNP). Estas partculas se mueven linealmente a travs de la canasta
nuclear. Al igual que las importinas, las RNP son recicladas hacia el ncleo. En el
citoplasma, las CRBP (del ingls, cytoplasmic RNA-binding proteins) reemplazan a las
RNP para guiar a los ARNs a sus destinos citoslicos correctos.
Modelo de mecanismo de exportacin de ARNm a travs del CPN
Instituto Hernando Arias de Saavedra
BIOLOGA
13
C R O M A T I N A
La cromatina est formada por el ADN ms sus protenas asociadas, principalmente
histonas.
Morfolgicamente se distinguen dos tipos de cromatina en el nucleoplasma:
La eucromatina tiene un aspecto claro y est poco condensada, se cree que se produce
una mayor expresin gnica. La heterocromatina tiene un aspecto oscuro y est muy
condensada, se supone que hay una menor expresin gnica.
El nucleoplasma est rodeado por la envuelta nuclear y en l se encuentra la cromatina
que se puede considerar como el ADN (cido desoxirribonucleico) ms todas las
molculas relacionadas con su organizacin.
El ADN est formado por 4 desoxinucletidos (abreviado como nucletidos). Cada
nucletido contiene una sucesin de tres componentes: base, pentosa y grupo fosfato.
Las bases son cuatro, dos pricas: adenina (A) y guanina (G), y dos pirimidnicas:
timidina o Timina (T) y citosina (C).
La pentosa es la desoxiribosa.
Cada base se une a una pentosa formando un desoxinuclesido. Cada desoxinuclesido
se une un grupo fostato en un carbono de la pentosa formndose un desoxinucletido.
As, una cadena de ADN est formado por una sucesin de nucletidos unidos entre s
por los grupos fosfato. Esto es una cadena simple pero el ADN est formado por dos
cadenas simples gracias a la complementariedad que existe entre las bases A y T y entre
G y C, las cuales establecen uniones del tipo puentes de hidrgeno.
Las dos hebras son antiparalelas, es decir, que en los extremos tenemos el carbono 3' de
una cadena y el 5' de la otra y organizadas en forma de doble hlice de unos 2.5 nm de
anchura.
Los nucletidos no slo estn en el ADN. Pueden estar formando parte de otras
molculas con funciones totalmente diferentes. Por ejemplo el ATP (adenn trifosfato) es
la molcula de transferencia energtica, al AMPc (adenn monofostato cclico) que es un
segundo mensajero muy importante.
El ADN no se encuentra libre en el ncleo sino asociado a protenas como las histonas y
a todas aquellas protenas implicadas en su procesamiento, formando la cromatina.
Instituto Hernando Arias de Saavedra
BIOLOGA
14
Las histonas son protenas asociadas al ADN que determinan su organizacin.
Hay dos tipos: las nucleosmicas que son cuatro (H2A, H2B, H3 y H4) y la histona H1.
Las cuatro histonas nucleosmicas son las responsables de formar junto con el ADN los
denominados nucleosomas, que son la unidad estructural bsica de la cromatina.
En menor proporcin hay otras protenas que pueden estar asociadas al ADN. Entre ellas
se encuentran todas las protenas responsables de la sntesis y replicacin del ADN.
Tipos de cromatina
La cromatina (la sustancia que compone los ncleos de las clulas y que resulta de la
interaccin del ADN con las protenas histnicas, no histnicas y ARN) puede presentar
distintos grados de empaquetamiento o contraccin. Cuando los cromosomas se tien
con sustancias qumicas que se unen al ADN aparecen regiones densamente teidas y
regiones menos densamente teidas.
Cuando se observa al microscopio electrnico (ME) un ncleo de una clula en interfase
aparecen zonas claras y oscuras. Las oscuras se corresponden mayoritariamente con
cromatina compactada, denominada heterocromatina. La heterocromatina se suele
situar en las proximidades de la envuelta nuclear o en los alrededores del nuclolo. En las
zonas claras la cromatina se dispone de forma ms laxa y se denomina eucromatina. A
veces se observa una estructura ms o menos redondeada ms oscura que es el
nuclolo, una porcin de la cromatina relacionada con la produccin de ARN ribosmico
y ensamblaje de ribosomas, que veremos posteriormente.
La eucromatina suele corresponder a ADN que est transcribindose, mientras que la
heterocromatina es en su mayora transcripcionalmente inactiva.
La heterocromatina a su vez se divide en heterocromatina facultativa, es decir que puede
pasar de heterocromatina a eucromatina y viceversa y en heterocromatina constitutiva
que est siempre condensada y corresponde al 10-20 % de la heterocromatina. Aunque
existan porciones de esta heterocromatina que se transcribe normalmente aqu se
localizan porciones de cromatina que slo actuarn cuando la clula entra en mitosis o en
meiosis.
Existe un grado de compactacin mayor cuando la clula empaqueta la cromatina para
formar los cromosomas durante los procesos de divisin, bien en mitosis o en meiosis. En
los cromosomas se empaquetan tanto la heterocromatina como la eucromatina. Es
interesante sealar que cuando se ha producido la divisin celular y se vuelve a
desempaquetar a los cromosomas, la cromatina de cada uno de ellos suele ocupar un
territorio concreto dentro del interior nuclear.
Instituto Hernando Arias de Saavedra
BIOLOGA
15
Es decir, en el interior del ncleo no existe una madeja enredada de cromatina sino un
espacio compartimentalizado donde cada cromosoma suele formar un compartimento
ms o menos delimitado.
NIVELES DE ORGANIZACIN DE LA CROMATINA
Entonces:
La observacin a travs del microscopio ptico de un ncleo interfsico, nos permite
distinguir dos tipos de cromatina. La eucromatina o cromatina laxa, de localizacin
central, y la heterocromatina o cromatina densa, en la periferia del ncleo. La
heterocromatina representa aproximadamente el 10% del total de cromatina y es
considerada transcripcionalmente inactiva.
La eucromatina se encontrara al menos en dos estados, la eucromatina accesible, que
representa alrededor del 10%, donde se encuentran los genes que se estn
transcribiendo y la eucromatina poco accesible, ms condensada (pero menos que la
heterocromatina), donde estn los genes que la clula no est transcribiendo.
Si el ncleo celular se incuba con nucleasas, enzimas que digieren el ADN, las
secuencias que primero se digieren son las que portan los genes expresados por la
clula, lo que corrobora el menor grado de condensacin de la eucromatina.
Microfotografa electrnica del ncleo de un hepatocito. a. eucromatina; b. RER
Microfotografa electrnica del ncleo de un linfocito. a. eucromatina; b. heterocromatina, c. mitocondrial
Instituto Hernando Arias de Saavedra
BIOLOGA
16
H1 y formacin de la fibra de 10 nm
Diferencias entre eucromatina y heterocromatina
Diferencias genticas: los experimentos de construccin de mapas demuestran
que la mayor parte de los genes activos se localizan en la eucromatina. En los
ncleos interfsicos, la eucromatina se tie menos densamente debido al menor
grado de empaquetamiento, y en general se acepta que este es el estado ms
compatible con la actividad gnica y la transcripcin. Se han detectado muy pocos
genes activos en la heterocromatina. La principal diferencia entre la eucromatina y
la heterocromatina radica por tanto en la actividad de estos dos tipos de
cromatina.
Diferencias citolgicas: a nivel estructural, en los ncleos interfsicos, existe un
mayor grado de enrollamiento o empaquetamiento en la heterocromatina que en
la eucromatina.
Las caractersticas de la hetero y eucromatina son:
Caractersticas de la Cromatina
Tipo de cromatina Estado fsico Cambio qumico Tipo de genes Replicacin
Eucromatina Laxa Acetilada Activos Fase S temprana
Heterocromatina condensada Metilada Silentes Fase S tarda
Instituto Hernando Arias de Saavedra
BIOLOGA
17
Elementos diferenciados en la estructura cromosmica
La organizacin de la cromatina no es uniforme a lo largo de la estructura del
cromosoma. De hecho, se pueden distinguir una serie de elementos diferenciados:
los centrmeros (o constricciones primarias),
los telmeros (o extremos cromosmicos),
las regiones organizadoras del nuclolo (NORs segn la abreviatura en ingls) y
los crommeros, todos ellos caracterizados por contener secuencias especficas
de ADN.
Cuando el cromosoma en interfase se esparce artificialmente sobre agua, tiene la
apariencia de un collar de perlas. Las perlas son los nucleosomas, las unidades de
enrollamiento de la cromatina.
Los nucleosomas estn formados por un centro o "core" de histonas. Dicho centro
posee dos copias de cada una de las siguientes histonas: H2A; H2B; H3 y H4
Alrededor del centro de histonas, 146 pares de bases del ADN se enrollan en dos vueltas.
La unin de las histonas al ADN no depende de una secuencia particular de nucletidos,
sino de la secuencia de aminocidos de la histona. Las histonas son unas de las
molculas ms conservadas durante el transcurso de la evolucin. La histona H4 en el
ternero difiere de la H4 de la planta de poroto en slo 2 residuos de aminocidos de una
cadena de 102. Alrededor de 60 pares de bases de ADN unen un nucleosoma con el
prximo. Cada regin de unin es el ADN espaciador.
La quinta histona, la H1, conecta a los nucleosomas y acta como una banda de goma,
mantenindolos juntos dentro de una misma cuerda enrollada. Esta estructura se conoce
como fibra de 10nm, siendo el primer grado del empaquetamiento de la cromatina.
Instituto Hernando Arias de Saavedra
BIOLOGA
18
Los nucleosomas se organizan, a su vez, en fibras de 30nm (solenoide), girando a
manera de resorte alrededor de un eje virtual. Esta estructura es mantenida por la
interaccin de las H1 de nucleosomas cercanos.
En el siguiente nivel de empaquetamiento, las fibras de 30 nm se organizan en una serie
de bucles o asas superenrolladas. Estos bucles se estabilizan gracias a la interaccin con
las protenas de la matriz nuclear o andamiaje nuclear (scaffold).
Cada bucle de cromatina representa un dominio funcional o unidad de replicacin. Estos
dominios contienen alrededor de 100.000 pares de bases, extensin de ADN suficiente
para acomodar varios genes de tamao promedio. Algunos genes, sin embargo, pueden
abarcar varios dominios adyacentes de un cromosoma. Cada cromosoma puede tener
cien o ms dominios. Durante la profase, los cromosomas aparecen en forma ms
condensada, alcanzando la cromatina su mayor nivel de condensacin en metafase. La
organizacin de los cromosomas envuelve la fosforilacin de la H1 y otras protenas, lo
cual causa el plegamiento y empaquetamiento an ms compacto de la cromatina. El
andamiaje o matriz nuclear se convierte en el centro de la estructura del cromosoma, y
como la compactacin contina, ste se pliega modo de acorden.
Empaquetamiento de la fibra de 30 nm
Instituto Hernando Arias de Saavedra
BIOLOGA
19
Modelo de empaquetamiento de la cromatina
Con coloraciones especiales los cromosomas, revelan diferencias estructurales de
importancia funcional. Las bandas oscuras consisten en cromatina altamente
condensada, mientras que las bandas claras se corresponden con cromatina ms laxa.
El examen de la cromatina en bandas claras y oscuras revela que ambos tipos de
cromatina estn acomodados en bucles de distintos tamaos y que a su vez se proyectan
desde el andamiaje plegado. El andamiaje est muy plegado en la heterocromatina, y es
ms lineal en las bandas de eucromatina, formando bucles ms amplios. Las histonas de
la eucromatina estn fuertemente acetiladas. Estos cambios afectaran el grado de
empaquetamiento de la eucromatina, hacindola ms accesible para la transcripcin de
sus genes.
Instituto Hernando Arias de Saavedra
BIOLOGA
20
El empaquetamiento de la cromatina permite confinar al ADN dentro del ncleo La
molcula de ADN de un cromosoma humano contiene 50 x 10
6
pares de nucletidos en el
cromosoma ms pequeo (1.7 cm con la molcula extendida) a 250 x 10
6
pares de
nucletidos en el ms largo (8.5 cm). Midiendo extremo con extremo el total de
cromosomas de una clula humana diploide, el ADN se extiende ms de 2 metros. El
empaquetamiento del ADN en forma de cromatina, no solamente le permite entrar dentro
de los lmites del ncleo, sino tambin lo protege del ataque de las nucleasas.
Estructura y composicin qumica de la cromatina
Los principales componentes que se obtienen cuando se asla la cromatina de los
ncleos interfsicos son el ADN, las protenas histnicas, las protenas no histnicas
y el ARN.
La cantidad de protenas no histnicas puede variar de unos tejidos a otros en el mismo
individuo y dentro del mismo tejido a lo largo del desarrollo.
Las histonas
Las histonas son protenas bsicas, ricas en residuos de lisina y arginina, que muestran
una elevada conservacin evolutiva y que interaccionan con el ADN formando una
subunidad que se repite a lo largo de la cromatina denominada nucleosoma.
Los principales tipos de histonas que se han aislado en los ncleos interfsicos en
diferentes especies eucariontes son: H1, H2A, H2B, H3 y H4.
Adems de estas histonas, tambin existen otras que son especficas de tejido como la
histona H5 muy rica en lisina, especfica de eritrocitos nucleados de vertebrados no
mamferos, y las histonas del endosperma. Asimismo, la cromatina centromrica se
caracteriza por la presencia de una isoforma especfica de la histona H3, denominada
CENP-A en vertebrados.
Una de las caractersticas ms destacables es su elevado conservadurismo evolutivo,
sobre todo de las histonas H3 y H4. La histona H4 de guisante y de timo de ternera, se
diferencian solamente en dos aminocidos. Este dato indica que las interacciones entre el
ADN y las histonas para formar la cromatina deben ser muy semejantes en todos los
organismos eucariontes.
Los genes que codifican las histonas se encuentran agrupados en nichos (o clusters)
que se repiten decenas o centenas de veces. Cada cluster o grupo contiene el siguiente
orden de genes que codifican histonas: H1-H2A-H3-H2B-H4. Estos genes son ricos en
pares G-C, ya que codifican protenas con un elevado contenido en lisina y arginina, pero
estn separados por secuencias espaciadoras ricas en pares A-T.
Instituto Hernando Arias de Saavedra
BIOLOGA
21
El nucleosoma
La cromatina de ncleos en interfase, cuando se observa mediante tcnicas de
microscopia electrnica, se puede describir como un collar de cuentas o un rosario, en el
que cada cuenta es una subunidad esfrica o globular que se denomina nucleosoma; los
nucleosomas se hallan unidos entre s mediante fibras de ADN. Se sigue, entonces, que
la unidad bsica de la estructura de la cromatina es el nucleosoma.
Un nucleosoma tpico est asociado a 200 pares de bases (pb) de ADN y est formado
por una mdula (core en ingls) y un ligador (o linker). La mdula est formada por un
octmero constituido por dos subunidades de las histonas H2A, H2B, H3 y H4.
El resto del ADN (60 pb) forma parte del ligador (linker), que interacciona con la histona
H1. La cantidad de ADN asociado con un nucleosoma vara de una especie a otra, de
154 pb a 241 pb; esta variacin se debe fundamentalmente a la cantidad de ADN
asociada al ligador (linker).
Estructura del nucleosoma
Instituto Hernando Arias de Saavedra
BIOLOGA
22
Protenas cromosmicas no histnicas: el armazn proteico
Las protenas cromosmicas no histnicas son protenas diferentes de las histonas
tienen un alto contenido en aminocidos bsicos (25% o ms), alto contenido en
aminocidos cidos (20-30%), una elevada proporcin de prolina (7%), bajo contenido en
aminocidos hidrofbicos y una alta movilidad electrofortica.
Las protenas HMG
Estas protenas se agrupan en una superfamilia por sus similitudes fsicas y qumicas, y
porque todas ellas actan como elementos arquitectnicos que afectan mltiples
procesos dependientes de ADN en el contexto de la cromatina. Todas las HMGs tienen
un terminal carboxilo rico en aminocidos de tipo cido, y se clasifican en tres familias
(HMGA, HMGB y HMGN), cada una con un motivo funcional nico, que induce cambios
especficos en sus sitios de unin y participa en funciones celulares diferentes.
E L N U C L E O L O
El nuclolo es un compartimento nuclear formado por cromatina y visible al microscopio
ptico. Las clulas de mamferos contienen desde 1 a 5 nuclolos. Sus dimensiones
varan dependiendo de la actividad de la clula y pude llegar a ser muy grande, del orden
de micras de dimetro. Normalmente en las clulas que estn realizando una gran
sntesis proteica poseen nuclolos grandes. Durante la mitosis desaparece permitiendo la
reorganizacin de la cromatina que lo forma para constituir los cromosomas.
En el nuclolo tiene lugar la formacin de subunidades ribosmicas, la sntesis y
procesamiento de ARNr y actualmente se considera que desempea un importante papel
en la regulacin del ciclo celular.
El nuclolo es un aglomerado de fibras de cromatina de distintos cromosomas. En el
hombre, los pares 13,14, 15, 21 y 22, aportan sectores de cromatina que forman el
nuclolo. Todos estos cromosomas son acrocntricos y presentan constricciones
secundarias denominadas organizadores nucleolares (NOR), donde estn los genes que
codifican ARNr.
Entonces:
En el nuclolo se dan procesos relacionados con la generacin de los ribosomas: sntesis
y maduracin del ARN ribosmico (ARNr) y ensamblaje de las subunidades ribosmicas.
El ensamblaje de las subunidades ribosmicas es un proceso curioso de trasiego de
molculas entre el citoplasma y el nucleoplasma. Primero se transcriben los genes de
dichas protenas, que se localizan fuera de la cromatina nucleolar, a ARNm que debe salir
al citosol donde es traducido a protenas por los ribosomas libres. Estas protenas
Instituto Hernando Arias de Saavedra
BIOLOGA
23
entrarn en el ncleo y llegan hasta el nuclolo. Aqu se asocian con los ARNr para forma
las subunidades ribosmicas que debern ser exportadas de nuevo al citosol atravesando
otra vez los poros nucleares. As, la visibilidad del nuclolo se debe a que muchos genes
que producen ARNr se estn transcribiendo, a que hay muchas protenas implicadas en el
procesamiento de ese primer transcrito, a las protenas de las subunidades ribosmicas y
a aquellas protenas relacionadas con su ensamblaje. Se estima que hay unas 690
protenas diferentes asociadas de forma estable con el nuclolo.
Esquema de nuclolo indicando los bucles de los 10 cromosomas con los genes para el ARNr
Distintas partes del nucleolo. El centro fibrilar es la zona donde se encuentran las copias de los genes que
codifican para el ARNr, el componente fribrilar denso es donde se produce el transcrito primario del ARNr y el
componente granular es donde se ensamblan las protenas y los diferentes ARNt para formar las subunidades
ribosmicas.
Instituto Hernando Arias de Saavedra
BIOLOGA
24
Morfolgicamente el nucleolo contiene distintas regiones:
el centro fibrilar, donde se encuentran los genes para el ARNr,
el componente fibrilar denso que rodea al centro fibrilar, donde se produce la
transcripcin activa de los genes ARNr, y
el componente granular donde se ensamblan las subunidades ribosmicas.
Microfotografa electrnica del nuclolo. NE, Envoltura nuclear; NO. Organizador nucleolar; PG, Zona
granular; PF, Zona fibrilar
Repasamos:
El nuclolo aparece como una estructura simple carente de componente membranoso, en
la que diferenciamos dos regiones:
Una zona fibrilar central, formada por ADNribosmico y ARNr naciente
Un zona granular perifrica donde los grnulos estn formados por las
subunidades ribosmicas en proceso de ensamblado
La mayora de las protenas presentes en la clula estn representadas slo por una
copia del gen que la codifica. Este es el caso de la hemoglobina de la sangre o
mioglobina de los msculos. Las protenas son abundantes porque de una sola copia se
traducen numerosas protenas. Se pueden producir ms de 10000 protenas por cada
molcula de ARNm. Esto no puede ocurrir cuando el destino de un gen es el propio ARN.
Una clula eucariota puede tener multitud de ribosomas y todos ellos contienen molculas
de ARNr. Una sola copia de un gen sera muy difcil que diera lugar a toda esta cantidad
de ARNr. La estrategia de las clulas es tener esos genes muy repetidos. Las clulas
humanas contienen unas 200 copias de sus genes para el precursor de los ARNr. Estos
Instituto Hernando Arias de Saavedra
BIOLOGA
25
genes se encuentran repartidos en 5 cromosomas diferentes. Adems los promotores y
las ARN polimerasas I presentan una gran afinidad por ellos. Todos estos genes se
agrupan en el nuclolo. Para el caso del ARNr 5S, uno de los ARNr que forman el
ribosoma, existen unas 20000 copias del gen, que se encuentra fuera del nuclolo, y es
transcrito por la polimerasa tipo III.
El transcrito primario de ARNr tiene que cortarse y procesarse para formar otros tipos de
ARNr que formarn el ribosoma: ARNr 18 S, ARNr 28 S y ARNr 4.5 S. El ARNr 5S
proviene de otra regin nuclear. El proceso de maduracin de este ARNr pre-ribosmico
se lleva a cabo por multitud de molculas de ARN nucleares pequeos. En el nuclolo se
estn produciendo constantemente estas secuencias a la vez que se procesan. Las dos
subunidades ribosmicas tendrn RNAs diferentes: la mayor 5.8S, 28S y 5S, y la menor
18S.
Los nuclolos, al igual que la envoltura nuclear desaparecen en la mitosis y se
reorganizan alrededor de los segmentos de ARNr, que como su nombre lo indica, codifica
ARNr. Siendo el ARNr el ms abundante dentro de los tipos de ARN, existen mltiples
copias del gen que lo codifica. El genoma humano presenta alrededor de 200 copias del
gen para ARNr. Estos genes que promedian los 10.000 nucletidos se localizan en
tndem. Cada gen est separado por ADN espaciador y presenta asociado una molcula
de ARN polimerasa I. De cada enzima parten perpendicularmente los ARNr nacientes,
tomando la apariencia caracterstica de un rbol de navidad. Cada gen produce un
transcripto llamado ARNr 45S que ser luego procesado.
El tamao del nucleolo vara entre clulas y en la misma clula segn su actividad, pues
si bien la velocidad de transcripcin puede acelerarse, el ensamblado de las subunidades
ribosomales requiere de un tiempo ms o menos constante; es por ello que en los
nuclolos grandes observamos mayor proporcin de componente granular.
Microfotografa electrnica de los genes nucleolares (ADNr) durante la transcripcin. Observe como la
longitud de los transcriptos primarios aumenta a medida que nos alejamos del punto de inicio.
Instituto Hernando Arias de Saavedra
BIOLOGA
26
Cromosoma
Vista general de las clulas en un pice de raz de cebolla (Allium cepa), observado con 800 aumentos. (a)
Clula sin dividirse, obsrvese la red de cromatina y el nuclolo intensamente teido; (b) ncleos preparados
para la divisin celular, puede observarse que la cromatina se ha condensado; (c) Clulas en distintos
estadios de divisin mittica, se pueden observar que la cromatina se ha terminado de condensar y se han
formado los cromosomas.
En biologa, se denomina cromosoma (del griego
, - chroma, color y , - soma,
cuerpo o elemento) a cada uno de los pequeos
cuerpos en forma de bastoncillos en que se organiza
la cromatina del ncleo celular durante las divisiones
celulares (mitosis y meiosis).
Como vimos anteriormente, la cromatina es un
material microscpico que lleva la informacin
gentica de los organismos eucariotas y est
constituida por ADN asociado a protenas especiales
llamadas histonas. Este material se encuentra en el ncleo de las clulas eucariotas y se
visualiza como una maraa de hilos delgados. Cuando el ncleo celular comienza el
proceso de divisin (cariocinesis), esa maraa de hilos inicia un fenmeno de
condensacin progresivo que finaliza en la formacin de entidades discretas e
independientes: los cromosomas.
Por lo tanto:
Cromatina y cromosoma son dos aspectos morfolgicamente distintos de una misma
entidad celular.
Instituto Hernando Arias de Saavedra
BIOLOGA
27
Diagrama de un cromosoma eucaritico duplicado y condensado (en metafase mittica). (1) Cromtida, cada
una de las partes idnticas de un cromosoma luego de la duplicacin del ADN. (2) Centrmero, el lugar del
cromosoma en el cual ambas cromtidas se tocan. (3) Brazo corto. (4) Brazo largo.
Cuando se examinan con detalle durante la mitosis, se observa que los cromosomas
presentan una forma y un tamao caractersticos. Cada cromosoma tiene una regin
condensada, o constreida, llamada centrmero, que confiere la apariencia general de
cada cromosoma y que permite clasificarlos segn la posicin del centrmero a lo largo
del cromosoma.
Instituto Hernando Arias de Saavedra
BIOLOGA
28
Otra observacin que se puede realizar es que el nmero de cromosomas de los
individuos de la misma especie es constante. Esta cantidad de cromosomas se denomina
nmero diploide y se simboliza como 2n. Cuando se examina la longitud de tales
cromosomas y la situacin del centrmero surge el segundo rasgo general: para cada
cromosoma con una longitud y una posicin del centrmero determinada existe otro
cromosoma con rasgos idnticos, o sea, casi todos los cromosomas se encuentran
formando parejas. Los miembros de cada par se denominan cromosomas homlogos.
Mapa citogentico o cariograma de una nia antes de nacer, resultado de una amniocentesis.
En la figura se presentan todos los cromosomas mitticos de una nia, ordenados por
parejas de homlogos y por su longitud, lo que se denomina cariotipo. Puede observarse
que en ese cariotipo hay 46 cromosomas (o sea, 2n=46) que es el nmero cromosmico
de la especie humana. Se puede advertir, tambin, que cada cromosoma tiene una
estructura doble, con dos cromtidas hermanas que yacen paralelas entre s y unidas por
un nico centrmero.
Durante la mitosis las cromtidas hermanas, que son idnticas, se separan una de otra
hacia dos nuevas clulas. Las parejas de cromosomas homlogos que se observan en la
imagen tienen, adems, una semejanza gentica fundamental: presentan los mismos
genes situados en los mismos lugares a lo largo del cromosoma (tales lugares se
denominan locus o loci en plural). Esto indica que cada miembro del par de homlogos
lleva informacin gentica para las mismas caractersticas del organismo.
En organismos con reproduccin sexual, uno de los miembros del par de cromosomas
homlogos proviene de la madre (a travs del vulo) y el otro del padre (a travs del
espermatozoide). Por ello, y como consecuencia de la herencia biparental, cada
Instituto Hernando Arias de Saavedra
BIOLOGA
29
organismo diploide tiene dos copias de cada uno de los genes, cada una ubicada en uno
de los cromosomas homlogos. Una excepcin importante en el concepto de parejas de
cromosomas homlogos es que en muchas especies los miembros de una pareja, los
cromosomas que determinan el sexo o cromosomas sexuales, no tienen usualmente el
mismo tamao, igual situacin del centrmero, la misma proporcin entre los brazos o,
incluso, los mismos loci. El cromosoma Y (que determina el sexo masculino en humanos)
es de menor tamao y carece de la mayora de los loci que se encuentran en el
cromosoma X.
Al agrupar estas parejas de homlogos existe un par de cromosomas que es diferente
segn estemos estudiando una hembra o un macho. Estos dos cromosomas que pueden
ser identificados por su forma y tamao, como pertenecientes a uno de los dos sexos, se
denominan cromosomas sexuales, mientras que los restantes pares de cromosomas
homlogos, que son iguales en tamao y forma para ambos sexos de una misma
especie, se denominan autosomas.
Autosomas 22 pares
Cromosomas
Cromosomas sexuales 1 par
La representacin grfica de todos los cromosomas de un individuo en cuanto al nmero,
tamao y forma constituye el CARIOTIPO.
Instituto Hernando Arias de Saavedra
BIOLOGA
30
Los cromosomas sexuales se han denominado X e Y. En los mamferos, las clulas de
los individuos machos contienen un par XY y las clulas de las hembras por un par XX.
En la especie humana, cuya dotacin cromosmica es de 46 cromosomas, cada clula
somtica contiene 22 pares de autosomas ms un par XX si se trata de una mujer y 22
pares de autosomas y un par XY si se trata de un varn.
La determinacin sexual queda marcada en el momento de la fecundacin y viene fijada
por el tipo de gametos que se unen. Las mujeres slo producirn un tipo de vulo con 22
autosomas y un cromosoma sexual X, mientras que los varones formarn dos tipos
de espermatozoides, el 50% portadores de un cromosoma X y el 50% portadores de un
cromosoma Y.
Al ser la fecundacin producto del azar, un vulo puede unirse a cualquiera de los tipos
de espermatozoides que se han producido, por lo que en la mitad de los casos se
formaran hembras y en otro 50% se formaran machos.
Centrmeros
El centrmero es la constriccin primaria. Es la zona por la que el cromosoma
interacciona con las fibras del huso acromtico desde profase hasta anafase, tanto en
mitosis como en meiosis, y es responsable de realizar y regular los movimientos
cromosmicos que tienen lugar durante estas fases. Las estructuras centromricas que
interaccionan con las fibras del huso se denominan cinetocoros. Adems, el centrmero
contribuye a la nucleacin de la cohesin de las cromtidas hermanas. En la estructura
del centrmero intervienen tanto el ADN centromrico, que consta fundamentalmente de
heterocromatina constitutiva, como protenas centromricas.
Instituto Hernando Arias de Saavedra
BIOLOGA
31
Telmeros
La palabra telmero procede del griego telos, "final" y meros, "parte".
Los telmeros son los extremos de los cromosomas. Son regiones de ADN no
codificante, altamente repetitivas, cuya funcin principal es la estabilidad estructural de
los cromosomas en las clulas eucariotas, la divisin celular y el tiempo de vida de las
estirpes celulares.
Adems estn involucradas en enfermedades tan importantes como el cncer. En los
organismos procariotes, los cromosomas son circulares y no poseen telmeros.
Cromosomas humanos (en gris) y sus telmeros (en blanco).
Los telmeros fueron descubiertos por Hermann Joseph Muller durante la dcada de los
aos 30. Desde entonces, se ha avanzado mucho en el conocimiento de los telmeros,
gracias a las tcnicas de la gentica molecular.
Estructura externa de los cromosomas: nmero, forma y tamao
El estudio de la estructura externa de los cromosomas de cualquier especie eucaritica
consiste en analizar la forma, tamao y nmero de los cromosomas que posee. El mejor
momento para llevar a cabo dicho estudio suele ser aquel en el que los cromosomas han
alcanzado su mximo grado de contraccin y tienen sus bordes perfectamente definidos.
Dicho momento suele ser la metafase mittica. El estudio de la estructura externa de los
cromosomas culmina con la obtencin del cariotipo. Los cromosomas se pueden estudiar
en distintos momentos segn la especie y dependiendo de los objetivos planteados.
Algunas especies tienen cromosomas que se pueden observar con gran detalle en
interfase, tal es el caso de Drosophila melanogaster, que posee cromosomas politnicos
gigantes que se observan en las glndulas salivales de dicho insecto, y el de Chironomus
tentans, otro dptero. El cariotipo se confecciona usualmente despus de un apropiado
pre-tratamiento y tincin de las clulas, para hacer ms visibles los cromosomas
individuales. Al diagrama simplificado de los cromosomas metafsicos del cariotipo se lo
denomina idiograma, que se construye con el nmero genmico. Para realizar el
ordenamiento de los cromosomas tanto en cariotipos como idiogramas se debe tener en
cuenta el tamao cromosmico (ubicados de mayor a menor, con el brazo corto bc o "p"
Instituto Hernando Arias de Saavedra
BIOLOGA
32
hacia arriba y el brazo largo bl o "q" hacia abajo); posicin del centrmero
(generalmente alineados) y presencia de constricciones secundarias y satlites.
Constancia del nmero de cromosomas
Nmeros de cromosomas en
diferentes especies
Especie
Nmero de
cromosomas
Hormiga Myrmecia pilosula,
macho
1
Hormiga Myrmecia pilosula,
hembra
2
Mosca de la fruta (Drosophila
melanogaster)
8
Centeno (Secale cereale) 14
Caracol (Helix) 24
Gato (Felis silvestris catus) 38
Cerdo (Sus scrofa) 40
Ratn (Mus musculus) 40
Trigo (Triticum aestivum) 42
Rata (Rattus rattus) 42
Conejo (Oryctolagus cuniculus) 44
Liebre (Lepus europaeus) 46
Humano (Homo sapiens sapiens) 46
Chimpanc (Pan troglodytes) 48
Patata, Papa (Solanum
tuberosum)
48
Oveja (Ovis aries) 54
Vaca (Bos taurus) 60
Asno (Equus asinus) 62
Mula (Equus mulus) 63 (estril)
Caballo (Equus caballus) 64
Camello ( Camelus bactrianus) 74
Llama (Lama glama) 74
Perro (Canis lupus familiaris) 78
Gallina (Gallus gallus) 78
Paloma Columbia livia 80
Pez Carassius auratus 94
Mariposa ~380
Helecho Ophioglussum reticulatum 1260
Protozoario Aulacantha
scolymantha
1600
Instituto Hernando Arias de Saavedra
BIOLOGA
33
Usualmente las especies animales y vegetales tienen un nmero de cromosomas constante
y determinado que constituyen su cariotipo (ley de la constancia numrica de los
cromosomas), aunque existen especies con una alta variabilidad cariotpica, no slo en
nmero sino en forma y tamao de los cromosomas.
El protozoario Aulacantha, con 1600 cromosomas en sus clulas, es la
especie con el mayor nmero de cromosomas.
El nmero de cromosomas de una especie (o fase vital) diploide se identifica como 2n
mientras que ese nmero en una especie (o fase vital) haploide se identifica con la letra n.
En aquellas especies que presentan un nmero repetido de cromosomas superior a dos
complementos se habla de poliploida, representndose el mltiplo por delante de la letra n.
As: 3n indicara un complemento cromosmico triploide, 4n un tetraploide, etc. Todas
estas son situaciones de euploida. Con la indicacin x se quiere expresar el nmero bsico
de cromosomas de una especie que presenta individuos con diversos grados de ploida o el
de una lnea filogentica a partir de la cual diversos taxones han alcanzado situaciones
aneuploides variadas, siendo en este caso el nmero cromosmico una variacin del
nmero original con aumento o disminucin del nmero bsico, por prdida, fusin o divisin
de cromosomas (p. ej., n+1 o n-1). Un ejemplo de esta situacin anormal la tenemos en los
individuos de la especie humana que presentan el llamado sndrome de Down, situacin de
aneuploida (2n=47) por la presencia de un ejemplar ms de lo habitual del cromosoma 21
(trisoma).
El cromosoma en organismos procariotas
Los procariotas, bacteria y archaea, presentan tpicamente un solo cromosoma circular, si
bien existen algunas variantes a esta regla. El cromosoma bacteriano puede tener un
tamao desde 160.000 pares de bases (como en el endosimbionte Carsonella ruddii a
12.200.000 pares de bases en la bacteria del suelo Sorangium cellulosum.
Las bacterias usualmente tienen un solo punto en su cromosoma desde el cual se inicia la
duplicacin, mientras que algunas archeas presentan mltiples sitios de inicio de la
duplicacin. Por otro lado, los genes de los procariotas estn organizados en operones y no
contienen intrones.
Instituto Hernando Arias de Saavedra
BIOLOGA
34
Los procariotas no poseen un ncleo verdadero, en cambio su ADN est organizado en una
estructura denominada nucleoide.
El nucleoide es una estructura distintiva y ocupa una regin definida en la clula bacteriana.
Esta estructura es muy dinmica y se halla mantenida y remodelada a travs de la accin de
protenas similares a histonas, las cuales se asocian al cromosoma bacteriano. En archaea,
el ADN en el cromosoma se halla todava ms organizado, con el ADN empacado dentro de
estructuras similares a los nucleosomas eucariticos.
EL CROMOSOMA EUCARIOTA
Cada cromosoma eucariota consiste en una molcula simple de ADN de alrededor de 150
millones de pares de nucletidos.
La molcula de ADN en el cromosoma eucariota es lineal, por lo tanto posee dos extremos
(en contraste con el cromosoma bacteriano que es circular).
La molcula de ADN de un cromosoma tpico eucariota contiene:
Un conjunto lineal de genes que codifican para ARN y protenas interrumpido por
Muchas secuencias de ADN no codificante.
El ADN no codificante incluye:
Secuencias de aproximadamente 170 nucletidos de ADN satlite, repetidas miles
de veces, que corresponden al centrmero.
Secuencias repetitivas en los extremos del cromosoma llamadas telmeros.
Mltiples secuencias sealizadoras altamente conservadas, denominadas origen de
replicacin (ORI) , necesarias para que se realice la duplicacin del ADN en un
tiempo breve.
Secuencias de un cromosoma eucariota estable en las diferentes etapas del ciclo celular
El centrmero es una constriccin primaria localizada centralmente o hacia los extremos de
cada cromosoma.
Instituto Hernando Arias de Saavedra
BIOLOGA
35
El ADN centromrico como ya mencionamos es altamente repetitivo y se encuentra siempre
condensado siendo parte de la heterocromatina.
Sntesis de telmeros por la telomerasa. a) La telomerasa se une al telomero; b) La telomerasa alarga los
extremos de la cadena 3`; c) La telomerasa avanza; d) La ADN polimerasa sintetiza la cadena retrasada.
Los telmeros son cruciales en la vida de la clula. Ellos son necesarios para la duplicacin
completa del cromosoma, los protegen de las nucleasas, evitan que los extremos del
cromosoma se fusionen entre s y facilitan la interaccin del cromosoma con la envoltura
nuclear.
Los telmeros de las clulas humanas contienen la secuencia 5'TTAGGG3, que se repite
aproximadamente 2000 veces.
5 '... ..TTAGGG TTAGGG TTAGGG TTAGGG TTAGGG TTAGGG..... 3 '
3 '... ..AATCCC AATCCC AATCCC AATCCC A..... 5 '
La cadena rica en guanosina corre en direccin 5' a 3', extendindose 12 a 15 nucletidos
ms all de la cadena rica en citosina, formando un apndice en una de las cadenas en
cada extremo del cromosoma. Este desnivel se mantiene de generacin en generacin por
Instituto Hernando Arias de Saavedra
BIOLOGA
36
medio de una enzima especial, la telomerasa, que agrega nuevas unidades al extremo 3' de
la cadena rica en guanosina.
La telomerasa es una ribonucleoprotena, la cual provee un molde de AAUCCC que gua la
insercin de la secuencia TTAGGG. Entonces la telomerasa es una retrotranscriptasa,
sintetiza ADN a partir de un molde de ARN.
Las clulas con telomerasa activa pueden compensar el acortamiento de los telmeros
durante la duplicacin del ADN.
La telomerasa activa se encuentra solamente en:
Las clulas de la lnea germinal, incluyendo clulas troncales embrionarias
Eucariotas unicelulares
Clulas cancerosas
Los cromosomas se diferencian por la ubicacin del centrmero
Durante la mayor parte de la vida de la clula, los cromosomas son demasiado largos y
tenues para ser vistos bajo un microscopio.
Microfotografa electrnica de un cromosoma metafsico
Antes de que una clula se divida, cada cromosoma se duplica (durante la fase S del ciclo
celular).
Al inicio de la divisin celular, los cromosomas duplicados se condensan en estructuras que
pueden teirse con facilidad (por ello denominadas cromosomas), pudindose observar bajo
el MO.
La condensacin es tal que el cromosoma es aproximadamente 10.000 veces ms corto que
la molcula de ADN que contiene. A primera vista, los cromosomas duplicados se
mantienen juntos por el centrmero. Mientras estn juntos, es comn llamar a cada parte del
cromosoma duplicado, cromtida hermana.
Esto no debe confundirnos,
Cada una de las "cromtidas hermanas" es un cromosoma completo.
Instituto Hernando Arias de Saavedra
BIOLOGA
37
El cinetocoro es una estructura proteica discoidal que forma parte del centrmero y ayuda a
separar las cromtidas hermanas. Es el sitio de unin con los microtbulos del huso, que
contienen los motores de dinena que tiran a los cromosomas en la anafase. Adems
proveen una plataforma para ensamblar y movilizar las protenas que construyen el huso.
El cromosoma se encuentra constituido bsicamente por el centrmero que divide el
cromosoma en un brazo corto o brazo p y un brazo largo o brazo q. Algunos cromosomas
presentan satlites en el brazo corto.
La forma de los cromosomas es para todas las clulas somticas constante y
caracterstica de cada especie. La forma depende fundamentalmente de las constricciones
que presente el cromosoma y de su localizacin en la cromtida.
Forma de los cromosomas
La posicin del centrmero, determina el largo de los brazos del cromosoma; en base a esto
se puede clasificar a los cromosomas en:
Metacntricos: el centrmero en posicin central determina brazos de igual longitud
Submetacntricos: un par de brazos es ms corto que el otro, pues el centrmero
se encuentra alejado del centro.
Acrocntricos: el centrmero se halla prximo a uno de los extremos, por lo tanto
uno de los brazos es casi inexistente.
Telocntricos: Slo se aprecia un brazo del cromosoma al estar el centrmero en el
extremo.
Tipos de cromosomas
Los cromosomas acrocntricos poseen una masa de cromatina llamada satlite, en el
extremo del brazo corto. El satlite se halla aislado del resto del cromosoma por la
Instituto Hernando Arias de Saavedra
BIOLOGA
38
constriccin secundaria. La zona aledaa al satlite de los cromosomas acrocntricos
contribuye a formar el nuclolo.
El ms corto de los dos brazos del cromosoma se llama p; el ms largo es el brazo q.
Partes de un cromosoma mittico.
El cromosoma humano 19 es metacntrico. El cromosoma humano 14 es acrocntrico
El cariotipo es una representacin grfica o fotogrfica de los cromosomas presentes en el
ncleo de una sola clula somtica de un individuo. Cada miembro del par de cromosomas
homlogos proviene de cada uno de los padres del individuo cuyas clulas examinamos.
Todas las especies tienen un nmero caracterstico de pares de cromosomas homlogos
llamado nmero diploide (2n). El nmero diploide del hombre es 46.
Instituto Hernando Arias de Saavedra
BIOLOGA
39
El cariotipo de la mujer contiene 23 pares de cromosomas homlogos, 22 pares son
autosomas y el par restante, cromosomas sexuales, ambos " X".
Cromosomas sexuales
En muchos organismos, uno de los pares de los cromosomas homlogos es distinto al resto,
realizando la determinacin del sexo del individuo. A estos cromosomas se les llama
cromosomas sexuales o heterocromosomas e incluso gonosomas, porque determinan el
sexo.
Sistema de determinacin XY: es propio del ser humano y muchos otros animales.
Las hembras, siendo XX, darn gametos iguales con cromosoma X, sexo
homogamtico y los machos, siendo XY, darn dos tipos de gametos, uno con el
cromosoma X y otro con el cromosoma Y. La probabilidad de que en la fecundacin,
al unirse los gametos, resulte una combinacin XX (hembra) o XY (macho) es
aproximadamente del 50%.
Sistema de determinacin ZW: en otras especies (mariposas, p.e.) ocurre lo
contrario, el sexo masculino es homogamtico (ZZ) y el femenino heterogamtico
(ZW).
Sistema de determinacin XO: otras especies (peces, insectos, anfibios) que no
tienen el cromosoma Y, determinndose el sexo por el nmero de cromosomas X,
macho XO y hembra XX.
El cariotipo del hombre contiene los mismos 22 pares de autosomas y 1 par de cromosomas
sexuales, un cromosoma sexual "X" y un cromosoma sexual "Y" (un gen en el cromosoma Y
Instituto Hernando Arias de Saavedra
BIOLOGA
40
designado SRY es el que pone en marcha el desarrollo de un varn, por lo tanto determina
el sexo).
El anlisis del cariotipo involucra la comparacin de cromosomas por su longitud, la
ubicacin de los centrmeros y la ubicacin y los tamaos de las bandas G.
Durante la mitosis, los 23 pares de cromosomas humanos se condensan y son visibles con
un microscopio ptico.
Preparacin de un Cariotipo: La preparacin de un cariotipo normalmente involucra
bloquear las clulas (glbulos blancos) durante la mitosis con colchicina y marcar los
cromosomas condensados con tincin Giemsa. La tincin marca las regiones de los
cromosomas que son ricos en pares de nucletidos entre A -T produciendo una banda
oscura, la banda G. Luego de la tincin, los cromosomas se fotografan, se recortan y se
ordenan de acuerdo a su longitud. Los de igual tamao se aparean segn la ubicacin de su
centrmero.
Una error comn es suponer que cada banda representa un slo gen. En realidad las
bandas ms pequeas contienen ms de un milln de pares de nucletidos y
potencialmente cientos de genes. Por ejemplo, el tamao de una banda pequea es igual a
toda la informacin gentica de una bacteria.
El anlisis del cariotipo es una de muchas tcnicas que nos permiten investigar las miles de
enfermedades genticas que se pueden encontrar en los seres humanos.
Cariotipo femenino normal: Los autosomas se ordenan en grupos por tamao y posicin del centrmero.
Instituto Hernando Arias de Saavedra
BIOLOGA
41
Mapa estndar (Idiograma) de patrones de bandeo del cromosoma 2. Los nmeros corresponden a las regiones
y bandas. A la derecha, idiograma del cariotipo humano masculino.
Genoma humano
El ser humano presenta 23 pares de cromosomas en sus clulas somticas: 22
autosomas y un par de cromosomas sexuales (dos X en el caso de las mujeres y un
cromosoma X y un Y en el caso de los varones). El tamao total aproximado del genoma
humano es de 3200 millones de pares de bases de ADN (3200 Mb) que contienen unos
20.000-25.000 genes. De las 3200 Mb unas 2950 Mb corresponden a eucromatina y unas
250 Mb a heterocromatina. El Proyecto Genoma Humano produjo una secuencia de
referencia del genoma humano eucromtico, usado en todo el mundo en las ciencias
biomdicas.
La secuencia de ADN que conforma el genoma humano contiene codificada la informacin
necesaria para la expresin, altamente coordinada y adaptable al ambiente, del proteoma
humano, es decir, del conjunto de protenas del ser humano.
Instituto Hernando Arias de Saavedra
BIOLOGA
42
REPRODUCCIN
La funcin de reproduccin. Es la generacin de nuevos individuos. Hay dos tipos de
reproduccin, la reproduccin asexual y la reproduccin sexual.
La reproduccin asexual. Es aquella en la que los descendentes son genticamente
idnticos al progenitor, es decir tienen la misma informacin en su ADN. Un ejemplo de
reproduccin asexual es el de una rama de geranio que se rompe y se planta en tierra. Al
cabo de un tiempo la rama genera races y se forma un nuevo geranio. En la reproduccin
asexual slo hay un progenitor y un proceso de multiplicacin celular en el cual las clulas
hijas son idnticas a la clula madre. Este tipo de divisin celular se denomina mitosis.
Tipo de reproduccin asexual en los organismos unicelulares. Segn la forma de
dividirse la clula se distingue la biparticin, la gemacin y la esporulacin.
Biparticin
Gemacin
Esporulacin
Tipo de reproduccin asexual en los organismos pluricelulares. Bsicamente consiste
en un fragmento del progenitor que crece y da lugar a un nuevo individuo. Se distingue la
reproduccin por esquejes en el geranio, por tubrculos en la patata, por bulbos en la
cebolla y por escisin o por gemacin en los plipos.
Instituto Hernando Arias de Saavedra
BIOLOGA
43
La reproduccin sexual. Es aquella en la que los descendentes son genticamente
diferentes de sus progenitores y diferente tambin entre los hermanos. Se realiza mediante
clulas especiales denominadas clulas sexuales que slo tienen la mitad de informacin
gentica y que es diferente en cada una de ellas.
Las clulas sexuales se originan mediante una divisin celular especial denominada
meiosis. Hay dos tipos de clulas sexuales: los gametos y las esporas sexuales.
Reproduccin sexual por gametos. Se realiza mediante la unin (fecundacin) de un
gameto masculino con un gameto femenino. Esto da lugar a una clula (zigoto) que ya tiene
la informacin gentica completa.
El zigoto por multiplicacin da lugar a un embrin y despus a todo un nuevo individuo.
Los gametos masculinos de los animales se denominan espermatozoides y los de las
plantas anterozoides. Los gametos femeninos de los animales se llaman vulos y los de
las plantas oosferas.
La fecundacin puede ser externa o interna gracias a rganos copuladores. En los
animales el desarrollo embrionario se puede producir dentro de un huevo (ovparos) o en el
interior del cuerpo materno (vivparos).
Reproduccin mediante esporas sexuales. En ella una sola espora ya genera todo un
nuevo individuo. Se da en hongos y en plantas. En estas ltimas se alterna una
reproduccin sexual mediante gametos con una reproduccin sexual mediante esporas.
Reproduccin alternante. Se da por ejemplo en algunas especies de medusas En ella se
alterna una reproduccin sexual por gametos con una reproduccin asexual mediante
fragmentacin
Fisin binaria
Instituto Hernando Arias de Saavedra
BIOLOGA
44
Fisin binaria.
La fisin binaria o biparticin es una forma de reproduccin asexual que se lleva a cabo
en bacterias, levaduras de fisin, algas unicelulares y protozoos. La clula madre se divide
en dos clulas hijas de igual tamao.
Efectos genticos
La mayor parte de las bacterias se reproducen por fisin binaria, lo que produce una tasa de
crecimiento exponencial. Por ejemplo, bajo condiciones ptimas, la bacteria Escherichia coli
se puede dividir una vez cada 20 minutos. El ADN bacteriano tiene tasas de mutacin
elevadas. De esta manera, la rpida reproduccin bacteriana da amplias oportunidades para
que se produzcan nuevas cepas capaces de desarrollar resistencia a antibiticos y les
ayuda a proliferar en una gran variedad de ambientes.
La fisin binaria comienza con la replicacin del ADN que en procariotas consta de una sola
molcula circular. Esta tiene lugar desde el origen de replicacin, que se abre formando una
burbuja de replicacin que separa el ADN doble hebra. Este nuevo ADN se va a anclar a la
membrana plasmtica en los polos de la clula a travs de desmosomas.
Instituto Hernando Arias de Saavedra
BIOLOGA
45
La evolucin de los organismos eucariotas y la creciente complejidad, tamao y nmero de
sus cromosomas hizo que se desarrollaran mecanismos ms elaborados para repartir el
material gnico en partes iguales a las clulas hijas. Esto dio lugar al aparato mittico que
en el caso de algunos eucariotas primitivos, como el dinoflagelado
2
(Crypthecodinium cohnii)
se da una situacin intermedia. En este organismo ocurre el anclaje de cromosomas a la
membrana plasmtica, una membrana nuclear que no se desintegra, microtbulos del huso
que se mantienen fuera del ncleo que donde presionan forman canales paralelos que
dividen al ncleo. Los cromosomas se dividen de forma igualitaria porque se anclan en los
polos opuestos de la membrana nuclear.
Tipos de fisin binaria
La fisin binaria puede dividirse en diferentes grupos dependiendo del plano de divisin:
Regular: una clula se divide simtricamente en dos partes de igual tamao
Tipo ameba: La divisin es un tanto irregular con respecto al citoplasma y
perpendicular respecto al eje del huso. Divisiones de este tipo, tienen lugar en
rizpodos.
Longitudinal: El eje de la divisin es longitudinal. Los flagelados poseen divisiones de
este tipo.
Transversal: Ocurre en ciliados como el Paramecium, el citoplasma se divide de
forma perpendicular al eje del huso.
2
Dinoflagellata o Pyrrhophyta es un extenso grupo de protistas flagelados. Son unicelulares y forman parte del
plancton marino, si bien las hay de agua dulce y coloniales. Sus poblaciones se distribuyen en funcin de la
temperatura, salinidad y profundidad. Alrededor de la mitad de los dinoflagelados son fotosintticos y constituyen
el grupo ms grande de algas eucariontes aparte de las Diatomeas. Puesto que son productores primarios,
constituyen una parte importante de la cadena alimenticia acutica. Ciertas especies fotosintticas, las
zooxantelas, son endosimbiontes de animales (corales, anmonas y almejas) y protozoos marinos
desempeando un papel importante en la biologa de los arrecifes coralinos. Otros dinoflagelados son
depredadores de otros protozoos y algunas formas son parsitas (por ejemplo, Oodinium y Pfiesteria). Algunos
dinoflagelados son responsables de las mareas rojas, pues sintetizan fuertes toxinas. El nombre procede del
griego dinos, girando y del latn, flagellum, ltigo.
Estructura de un dinoflagelado: 1-placa de la teca, 2-cngulo con el flagelo transversal, 3-
sulcus con el flagelo longitudinal.
Instituto Hernando Arias de Saavedra
BIOLOGA
46
Oblicua: Sucede en opalinidas, que poseen filas oblicuas de cilios. La divisin
comienza siendo longitudinal pero luego se vuelve paralela a estas filas de cilios. Es
intermedia entre la longitudinal y la transversal.
Conjugacin bacteriana
Esquema de la conjugacin bacteriana. 1-La clula donante genera un pilus
3
. 2-El pilus se une a la clula
receptora y ambas clulas se aproximan. 3-El plsmido mvil se desarma y una de las cadenas de ADN es
transferida a la clula receptora. 4-Ambas clulas sintetizan la segunda cadena y regeneran un plsmido
completo. Adems, ambas clulas generan nuevos pili y son ahora viables como donantes.
3
En bacteriologa, los pili (singular pilus, del latn cabello) son estructuras en forma de pelo, ms cortas y finos
que los flagelos que se encuentran en la superficie de muchas bacterias. Los pilus corresponden a
evaginaciones de la membrana citoplasmtica a travs de los poros de la pared celular y la cpsula que asoman
al exterior. Los trminos fimbria y pilus son a menudo intercambiables, pero fimbria se suele reservar para los
pelos cortos que utilizan las bacterias para adherirse a las superficies, en tanto que pilus suele referir a los pelos
ligeramente ms largos que se utilizan en la conjugacin bacteriana para transferir material gentico. Algunas
bacterias usan los pili para el movimiento.
Instituto Hernando Arias de Saavedra
BIOLOGA
47
La conjugacin bacteriana es el proceso de transferencia de informacin gentica desde
una clula donadora a otra receptora. Este proceso fue descubierto por Joshua Lederberg y
Edward Tatum en 1946. Es frecuentemente considerado como el equivalente bacteriano del
sexo, ya que implica intercambio de informacin gentica entre dos individuos. Este proceso
es promovido por determinados tipos de plsmidos, que portan un conjunto de genes cuyos
productos participan en el proceso, y que requiere contactos directos entre ambas clulas,
con intervencin de estructuras superficiales especializadas y de funciones especficas
(pilus o pilis sexuales en los Gram negativos, y contacto ntimo en los Gram positivos).
Algunos de estos plsmidos se comportan como episomas, es decir, que pueden integrarse
en el cromosoma; en este caso, si se produce la conjugacin, se puede transferir el propio
plsmido ms un segmento adyacente del cromosoma, que a su vez podr recombinarse
con secuencias homlogas del cromosoma del receptor, dando lugar a un cromosoma
hbrido.
Ciclo celular
Ciclo celular.
El ciclo celular es un conjunto ordenado de sucesos que conducen al crecimiento de la
clula y la divisin en dos clulas hijas. Las clulas que no estn en divisin no se
consideran que estn en el ciclo celular. Las etapas, mostradas a la derecha, son G
1
-S-G
2
y
M.
El estado G
1
quiere decir "GAP 1"(Intervalo 1). El estado S representa "Sntesis". Este es el
estado cuando ocurre la replicacin del ADN. El estado G
2
representa "GAP 2"(Intervalo 2).
Instituto Hernando Arias de Saavedra
BIOLOGA
48
El estado M representa la fase M, y agrupa a la mitosis (reparto de material gentico
nuclear) y citocinesis (divisin del citoplasma). Las clulas que se encuentran en el ciclo
celular se denominan proliferantes y las que se encuentran en fase G
0
se llaman clulas
quiescentes. Todas las clulas se originan nicamente de otra existente con anterioridad. El
ciclo celular se inicia en el instante en que aparece una nueva clula, descendiente de otra
que se divide, y termina en el momento en que dicha clula, por divisin subsiguiente,
origina dos nuevas clulas hijas.
Instituto Hernando Arias de Saavedra
BIOLOGA
49
Comparacin entre la fisin binaria, mitosis y meiosis, tres tipos de divisin celular.
Fases del ciclo celular
La clula puede encontrarse en dos estados claramente diferenciados:
El estado de divisin, llamado fase M.
El estado de no divisin o interfase. La clula realiza sus funciones especficas y, si
est destinada a avanzar a la divisin celular, comienza por realizar la duplicacin de
su ADN.
Micrografas de: a la izquierda, interfase celular; despus, las distintas fases de la mitosis, dentro de la fase M del
ciclo celular.
Instituto Hernando Arias de Saavedra
BIOLOGA
50
Interfase
Es el perodo comprendido entre divisiones celulares. Es la fase ms larga del ciclo celular,
ocupando casi el 95% del ciclo, trascurre entre dos mitosis y comprende tres etapas:
Fase G
1
(del ingls Growth o Gap 1): Es la primera fase del ciclo celular, en la que
existe crecimiento celular con sntesis de protenas y de ARN. Es el perodo que
trascurre entre el fin de una mitosis y el inicio de la sntesis de ADN. Tiene una
duracin de entre 6 y 12 horas, y durante este tiempo la clula duplica su tamao y
masa debido a la continua sntesis de todos sus componentes, como resultado de la
expresin de los genes que codifican las protenas responsables de su fenotipo
particular. En cuanto a carga gentica, en humanos (diploides) son 2n 2c.
Fase S (del ingls Synthesis): Es la segunda fase del ciclo, en la que se produce la
replicacin o sntesis del ADN, como resultado cada cromosoma se duplica y
queda formado por dos cromtidas idnticas. Con la duplicacin del ADN, el ncleo
contiene el doble de protenas nucleares y de ADN que al principio. Tiene una
duracin de unos 6-8 horas.
Fase G
2
(del ingls Growth o Gap 2): Es la tercera fase de crecimiento del ciclo
celular en la que contina la sntesis de protenas y ARN. Al final de este perodo se
observa al microscopio cambios en la estructura celular, que indican el principio de la
divisin celular. Tiene una duracin entre 3 y 4 horas. Termina cuando la cromatina
empieza a condensarse al inicio de la mitosis. La carga gentica de humanos es 2n
4c, ya que se han duplicado los cromosomas, teniendo ahora dos cromtidas cada
uno.
Componentes reguladores
El ciclo celular es controlado por un sistema que vigila cada paso realizado. En regiones
concretas del ciclo, la clula comprueba que se cumplan las condiciones para pasar a la
etapa siguiente: de este modo, si no se cumplen estas condiciones, el ciclo se detiene.
Existen cuatro transiciones principales:
Paso de G
0
a G
1
: comienzo de la proliferacin.
Transicin de G
1
a S: iniciacin de la replicacin.
Paso de G
2
a M: iniciacin de la mitosis.
Avance de metafase a anafase
Los genes que regulan el ciclo celular se dividen en tres grandes grupos:
1. Genes que codifican protenas para el ciclo: enzimas y precursores de la sntesis de
ADN, enzimas para la sntesis y ensamblaje de tubulina, etc.
2. Genes que codifican protenas que regulan positivamente el ciclo: tambin llamados
protooncogenes. Las protenas que codifican activan la proliferacin celular, para que
Instituto Hernando Arias de Saavedra
BIOLOGA
51
clulas quiescentes pasen a la fase S y entren en divisin. Algunos de estos genes
codifican las protenas del sistema de ciclinas y quinasas dependientes de ciclina.
Pueden ser:
o Genes de respuesta temprana, inducidos a los 15 minutos del tratamiento con
factores de crecimiento, sin necesidad de sntesis proteica;
o Genes de respuesta tarda, inducidos ms de una hora despus del
tratamiento con factores de crecimiento, su induccin parece estar causada
por las protenas producidas por los genes de respuesta temprana.
3. Genes que codifican protenas que regulan negativamente el ciclo: Tambin
llamados genes supresores tumorales.
Las ciclinas y las quinasas dependientes de ciclina (CDK), son sintetizadas a partir de
protooncogenes y trabajan en cooperacin para regular el ciclo positivamente. Fosforilan
serinas y treoninas de protenas diana para desencadenar procesos celulares.
Los protooncogenes son genes cuya presencia o activacin a oncogenes pueden estimular
el desarrollo de cncer. Cuando se activan exageradamente en las clulas normales
provocan que ellas pierdan el control de la divisin y se mantengan proliferando sin control.
Expresin diferencial de ciclinas en las distintas fases del ciclo.
Las ciclinas son un grupo heterogneo de protenas que se distinguen segn el momento
del ciclo en el que actan. Las ciclinas son protenas de vida muy corta: tras disociarse de
sus kinasas asociadas, se degradan con extrema rapidez.
Las kinasas dependientes de ciclinas (CDK por sus siglas en ingls) son molculas de
mediano peso molecular que presentan una estructura proteica caracterstica, consistente
en dos lbulos entre los cuales est el centro cataltico, donde se inserta el ATP (que ser el
donador de grupos fosfato.
El proceso de sntesis y ensamblaje de ciclinas/CDK est regulado por tres tipos de factores:
mitgenos, que estimulan la divisin celular; factores de crecimiento (GFs), que producen un
Instituto Hernando Arias de Saavedra
BIOLOGA
52
aumento de tamao al estimular la sntesis proteica; y factores de supervivencia, que
suprimen la apoptosis
4
.
La verificacin se lleva a cabo en los puntos de control y asegura la fidelidad de la
replicacin y segregacin del genoma. Algunos componentes, adems de detectar fallos,
pueden poner en marcha la reparacin.
Regulacin del ciclo celular
Esquema global de los elementos ms relevantes implicados en la regulacin del ciclo celular.
La regulacin del ciclo celular, explicada en el ao 2001 en organismos eucariotas, puede
contemplarse desde la perspectiva de la toma de decisiones en puntos crticos,
especialmente en la mitosis. De este modo, se plantean algunas preguntas:
Cmo se replica el ADN una nica vez? Una pregunta interesante es cmo se
mantiene la euploida
5
celular. Sucede que, en la fase G
1
, la Cdk (ciclina) promueve
4
La apoptosis es una forma de muerte celular, que est regulada genticamente.
5
Euploida es el estado celular en el cual la clula tiene uno o ms juegos completos de cromosomas
(dotaciones monoploides (x)) de su especie; dependiendo de especies, se excluyen los cromosomas sexuales.
Los individuos con euploida pueden presentar o no un nmero anormal de cromosomas diferente al habitual. Por
ejemplo, un humano, tiene 46 cromosomas, que es un nmero mltiplo de 23, sus nmero de cromosomas
monoploide (x) (que en el caso de los humanos coincide con el nmero de cromosomas haploide (n); x = n = 23),
por lo tanto es euploide (2x = 2n = 46). Un humano hipottico anormal, pero con juegos enteros de cromosomas,
Instituto Hernando Arias de Saavedra
BIOLOGA
53
la adicin al complejo de reconocimiento del origen de replicacin del ADN de unos
reguladores llamados Cdc6, los cuales reclutan a Mcm, formando un complejo
prerreplicativo del ADN, que recluta a la maquinaria de replicacin gentica. Una vez
que se inicia la fase S, la Cdk-S produce la disociacin de Cdc6 y su posterior
protelisis
6
, as como la exportacin al citosol de Mcm, con lo que el origen de
replicacin no puede, hasta el ciclo siguiente, reclutar un complejo prerreplicativo (las
degradaciones proteolticas siempre conllevan irreversibilidad, hasta que el ciclo
gire). Durante G
2
y M se mantiene la unicidad de la estructura de prerreplicacin,
hasta que, tras la mitosis, el nivel de actividad Cdk caiga y se permita la adicin de
Cdc6 y Mdm para el ciclo siguiente.
Cmo se separan las cromtidas hermanas? Ya en mitosis, tras la formacin del
huso acromtico y superacin del punto de restriccin de unin a cinetocoros, las
cromtidas han de eliminar su esqueleto de cohesinas, que las unen. Para ello, Cdk-
M favorece la activacin de APC, una ligasa de ubiquitina, por unin a Cdc20. Esta
APC ubiquitiniza y favorece la ulterior degradacin en el proteasoma de la segurina,
inhibidor del enzima separasa que debe escindir las cohesinas.
Cmo se entra en mitosis? La ciclina B, tpica en la Cdk-M, existe en todo el ciclo
celular. Sucede que la Cdk(ciclina) est habitualmente inhibida por fosforilacin
mediante la protena Wee, pero, a finales de G2, se activa una fosfatasa llamada
Cdc25 que elimina el fosfato inhibidor y permite el aumento de su actividad. Cdk-M
inhibe a Wee y activa a Cdc25, lo que produce una retroalimentacin positiva que
permite la acumulacin de Cdk-M.
Cmo se sale de mitosis? Una vez que los niveles de Cdk-M son altos, parece difcil
detener la dinmica de mitosis y entrar en citocinesis: pues bien, esto ocurre porque
la APC activada por la Cdk-M, y tras un lapso cuyo mecanismo de control es an
desconocido, ubiquitiniza a la ciclina B, produciendo el cese absoluto de actividad
Cdk-M.
Cmo se mantiene el estado G1? En la fase G1, la actividad Cdk est muy
disminuida porque: APC-Hct1 (Cdc20 slo acta en mitosis) elimina toda ciclina B; se
acumulan inhibidores de Cdk; la transcripcin de ciclinas se ve disminuida. Para
por ejemplo, 69, sera tambin euploide, en tanto que 69 sera mltiplo entero del nmero monoploide (3x = 69).
No se debe confundir con la aneuploida, que, en contraposicin a la euploida, se refiere a una alteracin en la
cantidad de uno de los tipos de cromosomas homlogos. La euploida y el contenido de ADN de las clulas
eucariotas vara entre especies y entre las clulas de un mismo organismo. La euploida puede ser de dos tipos:
- Monoploida: la que poseen los organismos haploides (n); significa que solo tienen un nico cromosoma de
cada tipo.
- Poliploida: la poseen individuos con varios juegos completos de cromosomas homlogos (triploidas 3n,
tetraploidas 4n...).
6
La protelisis es la degradacin de protenas ya sea mediante enzimas especficas, llamadas proteasas, o
por medio de digestin intramolecular.
Instituto Hernando Arias de Saavedra
BIOLOGA
54
escapar de este reposo, se deben acumular ciclinas de G1. Esto se controla
mediante factores de proliferacin celular, seales externas. Los mecanismos
moleculares de activacin de transcripcin de genes de las fases S y G2 necesarios
para proseguir el ciclo son apasionantes: stos genes estn regulados por la protena
reguladora E2F, la cual se une a promotores de ciclinas G1/S y S. E2F est
controlada por la protena del retinoblastoma (Rb), la cual, en ausencia de factores
trficos, inhibe la actividad promotora de la transcripcin de E2F. Cuando existen
seales de proliferacin, Cdk-G1 fosforila Rb, que pierde afinidad por E2F, se disocia
de ste y permite que se expresen los genes de la fase S. Adems, como E2F
acelera la transcripcin de su propio gen, las Cdk-S y G1/S fosforilan tambin a Rb y
a Hct1 (activador de APC, que degradara estas ciclinas), se produce una
retroalimentacin positiva.
Ciclo celular en plantas
Los programas de desarrollo en plantas, a diferencia de lo que ocurre en animales, suceden
tras la embriognesis. La proliferacin y divisin celular est circunscrita a los meristemos
(tejidos responsables del crecimiento vegetal), zonas en las cuales se producen abundantes
divisiones celulares que dan lugar a la aparicin de nuevos rganos. Las hojas y las flores
derivan del meristemo apical del tallo y del meristemo floral, respectivamente, mientras que
el meristemo radicular da lugar a la raz. La regulacin, por tanto, de los programas de
desarrollo se basa en buena medida en la expresin gnica particular de los meristemos y
de la pauta concomitante de divisin celular; en plantas no existe la migracin celular como
mecanismo de desarrollo. La interaccin antagonstica entre las hormonas auxina
7
y
citoquinina
8
parece ser el mecanismo clave para el establecimiento de identidades y pautas
de proliferacin durante la embriognesis y durante el desarrollo de los meristemos caulinar
y radicular.
El ciclo celular de plantas comparte elementos comunes con el de animales, as como
ciertas particularidades. Las kinasas dependientes de ciclina (CDK) regulan, en buena
medida, las caractersticas del ciclo celular.
En cuanto a ciclinas, las plantas poseen una diversidad mayor que los animales: Arabidopsis
thaliana contiene como mnimo 32 ciclinas, quiz debido a los eventos de duplicacin de su
genoma. La expresin de las diferentes ciclinas parece estar regulada por diversas
fitohormonas.
7
Las auxinas son un grupo de fitohormonas que funcionan como reguladoras del crecimiento vegetal.
Esencialmente provocan la elongacin de las clulas. Se sintetizan en las regiones meristemticas del pice de
los tallos y se desplazan desde all hacia otras zonas de la planta, principalmente hacia la base, establecindose
as un gradiente de concentracin.
8
Las citoquininas o citocininas constituyen un grupo de hormonas vegetales que promueven la divisin y la
diferenciacin celular
Instituto Hernando Arias de Saavedra
BIOLOGA
55
Una caracterstica de gran flexibilidad de las clulas vegetales es la permisibilidad frente a
endorreduplicaciones, esto es, duplicaciones de la dotacin cromosmica (cambios de
ploida), que se deben a la replicacin del contenido gentico sin que medie una citocinesis.
Este mecanismo es usual en determinados tejidos pero tambin puede suceder en plantas
completas. Debido a que suele ir asociado a un mayor tamao celular, ha sido objeto de
seleccin en la mejora vegetal. Este hecho se explica debido al carcter ssil de los
organismos vegetales y, por tanto, la imposibilidad de ejecutar comportamientos de evitacin
frente a estreses ambientales; de este modo, las plantas estresadas con un mayor nmero
de copias del genoma podran ser ms resistentes. Los datos experimentales no siempre
apoyan esta hiptesis.
Fase M
Mitosis
Es la divisin celular en la que una clula progenitora (clulas eucariotas, clulas
somticas -clulas comunes del cuerpo-) se divide en dos clulas hijas idnticas. Esta
fase incluye la mitosis, a su vez dividida en: profase, metafase, anafase, telofase; y la
citocinesis, que se inicia ya en la telofase mittica. Si el ciclo completo durara 24 h, la
fase M durara alrededor de media hora (30 minutos).
Micrografa de una clula mittica pulmonar de tritn.
En biologa, la mitosis (del griego mitos, hebra) es un proceso de reparto equitativo del
material hereditario (ADN) caracterstico de las clulas eucariticas. Normalmente concluye
con la formacin de dos ncleos separados (cariocinesis), seguido de la particin del
citoplasma (citocinesis), para formar dos clulas hijas.
La mitosis completa, que produce clulas genticamente idnticas, es el fundamento del
crecimiento, de la reparacin tisular y de la reproduccin asexual. La meiosis, un proceso
que comparte mecanismos con la mitosis pero que no debe confundirse con ella (es otro tipo
de divisin celular, propio de los gametos), produce clulas genticamente distintas y,
combinada con la fecundacin, es el fundamento de la reproduccin sexual y la variabilidad
gentica.
Instituto Hernando Arias de Saavedra
BIOLOGA
56
La mitosis es el tipo de divisin celular por el cual se conservan las organelas y la
informacin gentica contenida en sus cromosomas, que pasa de esta manera a las clulas
hijas resultantes de la mitosis. La mitosis es igualmente un verdadero proceso de
multiplicacin celular que participa en el desarrollo, el crecimiento y la regeneracin del
organismo. Este proceso tiene lugar por medio de una serie de operaciones sucesivas que
se desarrollan de una manera continua, y que para facilitar su estudio han sido separadas
en varias etapas.
Esquema que muestra de manera resumida lo que ocurre durante la mitosis.
El resultado esencial de la mitosis es la continuidad de la informacin hereditaria de la clula
madre en cada una de las dos clulas hijas.
El genoma se compone de una determinada cantidad de genes organizados en
cromosomas, hebras de ADN muy enrolladas que contienen la informacin gentica vital
para la clula y el organismo. Dado que cada clula debe contener completa la informacin
gentica propia de su especie, la clula madre debe hacer una copia de cada cromosoma
antes de la mitosis, de forma que las dos clulas hijas reciban completa la informacin.
Esto ocurre durante la fase S de la interfase, el perodo que alterna con la mitosis en el ciclo
celular y en el que la clula entre otras cosas se prepara para dividirse.
Tras la duplicacin del ADN, cada cromosoma consistir en dos copias idnticas de la
misma hebra de ADN, llamadas cromtidas
9
hermanas, unidas entre s por una regin del
9
La cromtida es una de las unidades longitudinales de un cromosoma duplicado, unida a su cromtida
hermana por el centrmero. El cromonema es cada uno de los filamentos que componen la cromtida. Al
cromonema lo acompaan, a lo largo, una sucesin de grnulos a los que se ha dado el nombre de crommeros.
Instituto Hernando Arias de Saavedra
BIOLOGA
57
cromosoma llamada centrmero. Cada cromtida hermana no se considera en esa situacin
un cromosoma en s mismo, sino parte de un cromosoma que provisionalmente consta de
dos cromtidas.
1 Cromtida- 2 Centrmero- 3 Brazo Corto- 4 Brazo Largo.
En animales y plantas, pero no siempre en hongos o protistas, la envoltura nuclear que
separa el ADN del citoplasma se desintegra, desapareciendo la frontera que separaba el
contenido nuclear del citoplasma. Los cromosomas se ordenan en el plano ecuatorial de la
clula, perpendicular a un eje definido por un huso acromtico. ste es una estructura
citoesqueltica compleja, de forma ahusada, constituido por fibras que son filamentos de
microtbulos. Las fibras del huso dirigen el reparto de las cromtidas hermanas, una vez
producida su separacin, hacia los extremos del huso.
Por convenio cientfico, a partir de este momento cada cromtida hermana s se considera
un cromosoma completo, y empezamos a hablar de cromosomas hermanos para referirnos
a las estructuras idnticas que hasta ese momento llambamos cromtidas. Como la clula
se alarga, las fibras del huso tiran por el centrmero a los cromosomas hermanos
dirigindolos cada uno a uno de los polos de la clula. En las mitosis ms comunes,
llamadas abiertas, la envoltura nuclear se deshace al principio de la mitosis y se forman dos
envolturas nuevas sobre los dos grupos cromosmicos al acabar. En las mitosis cerradas,
que ocurren por ejemplo en levaduras, todo el reparto ocurre dentro del ncleo, que
finalmente se estrangula para formar dos ncleos separados.
Se llama cariocinesis a la formacin de los dos ncleos con que concluye habitualmente la
mitosis. Es posible, y ocurre en ciertos casos, que el reparto mittico se produzca sin
Est constituido por ADN y protenas. Los crommeros son un enrollamiento intenso del cromonema. Estn
unidos unos a otros a modo de cuentas de rosario. El conjunto de dos cromtidas generan un cromosoma. Cada
cromtida lleva varios alelos, es decir, cada una de las caractersticas de su progenitor. La cromtida es visible
en la metafase de la mitosis y representa a los pre-cromosomas hijos. En la anafase de la mitosis, las cromtidas
se separan y se convierten en cromosomas hijos. Son idnticas llevan la misma informacin, ya que se han
formado una a partir de la otra.
Instituto Hernando Arias de Saavedra
BIOLOGA
58
cariocinesis (endomitosis) dando lugar a un ncleo con el material hereditario duplicado
(doble nmero de cromosomas).
La mitosis se completa casi siempre con la llamada citocinesis o divisin del citoplasma. En
las clulas animales la citocinesis se realiza por estrangulacin: la clula se va estrechando
por el centro hasta que al final se separa en dos. En las clulas de las plantas se realiza por
tabicacin, es decir, las clulas hijas construyen una nueva regin de pared celular que
dividir la una de la otra dejando puentes de citoplasma (plasmodesmos). Al final, la clula
madre se parte por la mitad, dando lugar a dos clulas hijas, cada una con una copia
equivalente y completa del genoma original.
Cabe sealar que las clulas procariotas experimentan un proceso similar a la mitosis
llamado fisin binaria. No se puede considerar que las clulas procariotas experimenten
mitosis, dado que carecen de ncleo y nicamente tienen un cromosoma sin centrmero.
Fases del ciclo celular
La divisin de las clulas eucariotas es parte de un ciclo vital continuo, el ciclo celular, en el
que se distinguen dos perodos mayores, la interfase, durante la cual se produce la
duplicacin del ADN, y la mitosis, durante la cual se produce el reparto idntico del material
antes duplicado.
La mitosis es una fase relativamente corta en comparacin con la duracin de la interfase.
Interfase
La clula est ocupada en la actividad metablica preparndose para la mitosis (las
prximas cuatro fases que conducen e incluyen la divisin nuclear). Los cromosomas no se
disciernen claramente en el ncleo, aunque una mancha oscura llamada nuclolo, puede ser
visible. La clula puede contener un centrosoma con un par de centriolos (o centros de
organizacin de microtbulos en los vegetales) los cuales son sitios de organizacin para los
microtbulos.
Instituto Hernando Arias de Saavedra
BIOLOGA
59
Fases de la Mitosis
Diagrama mostrando los cambios que ocurren en los centrosomas y el ncleo de una clula en el proceso de la
divisin mittica. I a III, profase; IV, prometafase; V,metafase; VI y VII, anafase; VII y VIII, telofase.
Profase
Profase: Los dos centros de origen de los microtbulos (en verde) son los
centrosomas. La cromatina ha comenzado a condensarse y se observan las cromtidas (en azul). Las
estructuras en color rojo son los cinetocoros. (Micrografa obtenida utilizando marcajes fluorescentes).
Instituto Hernando Arias de Saavedra
BIOLOGA
60
Es la fase ms larga de la mitosis. Se produce en ella la condensacin del material
gentico (ADN, que en interfase existe en forma de cromatina), para formar unas
estructuras altamente organizadas, los cromosomas. Como el material gentico se ha
duplicado previamente durante la fase S, los cromosomas replicados estn formados por
dos cromtidas, unidas a travs del centrmero por molculas de cohesinas.
Uno de los hechos ms tempranos de la profase en las clulas animales es duplicacin del
centrosoma
10
. Los dos centrosomas hijos (cada uno con dos centriolos) migran entonces
hacia extremos opuestos de la clula. Los centrosomas actan como centros organizadores
de microtbulos, controlando la formacin de unas estructuras fibrosas, los microtbulos,
mediante la polimerizacin de tubulina soluble (protena globular). De esta forma, el huso de
una clula mittica tiene dos polos que emanan microtbulos.
En la profase tarda desaparece el nuclolo y se desorganiza la envoltura nuclear.
Papel del centrosoma en la progresin del ciclo celular
10
El centrosoma es un orgnulo celular que no est rodeado por una membrana; consiste de dos centriolos
apareados, embebidos en un conjunto de agregados proteicos que los rodean y que se denomina material
pericentriolar. Su funcin primaria consiste en la nucleacin y el anclaje de los microtbulos, por lo que de forma
genrica estas estructuras (conjuntamente con los cuerpos polares del huso en levaduras) se denominan centros
organizadores de microtbulos. Alrededor de los centrosomas se dispone radialmente un conjunto de
microtbulos formando un ster. Los centrosomas tienen un papel fundamental en el establecimiento de la red de
microtbulos en interfase y del huso mittico. Durante la mitosis, forman el huso mittico, necesario para la
segregacin de los cromosomas entre las dos clulas hija.
Instituto Hernando Arias de Saavedra
BIOLOGA
61
Prometafase
Prometafase: La membrana nuclear se ha disuelto, y los microtbulos (verde) invaden el espacio nuclear. Los
microtbulos pueden anclar cromosomas (azul) a travs de los cinetocoros (rojo) o interactuar con microtbulos
emanados por el polo opuesto.
La membrana nuclear se desensambla y los microtbulos invaden el espacio nuclear. Esto
se denomina mitosis abierta, y ocurre en una pequea parte de los organismos
multicelulares. Los hongos y algunos protistas, como las algas o las tricomonas11, realizan
una variacin denominada mitosis cerrada, en la que el huso se forma dentro del ncleo o
sus microtbulos pueden penetrar a travs de la membrana nuclear intacta.
Cada cromosoma ensambla dos cinetocoros hermanos sobre el centrmero, uno en cada
cromtida.
Un cinetocoro es una estructura proteica compleja a la que se anclan los microtbulos.
Aunque la estructura y la funcin del cinetocoro no se conocen completamente, contiene
varios motores moleculares, entre otros componentes. Cuando un microtbulo se ancla a
un cinetocoro, los motores se activan, utilizando energa de la hidrlisis del ATP para
"ascender" por el microtbulo hacia el centrosoma de origen. Esta actividad motora,
acoplada con la polimerizacin/despolimerizacin de los microtbulos, proporciona la fuerza
de empuje necesaria para separar ms adelante las dos cromtidas de los cromosomas.
Cuando el huso crece hasta una longitud suficiente, los microtbulos asociados a
cinetocoros empiezan a buscar cinetocoros a los que anclarse. Otros microtbulos no se
11
Trichomonas vaginalis es un protozoo patgeno flagelado perteneciente al orden Trichomonadida que
parasita el tracto urogenital tanto de hombres como de mujeres, pero nicamente en humanos. Produce una
patologa denominada tricomoniasis urogenital. Fue descrito por primera vez por Donn en 1836. Aos ms
tarde, en 1916, Hoehne demostr que este parsito era el responsable de un tipo de infeccin vaginal especfica.
Instituto Hernando Arias de Saavedra
BIOLOGA
62
asocian a cinetocoros, sino a otros microtbulos originados en el centrosoma opuesto para
formar el huso mittico. La prometafase se considera a veces como parte de la profase.
Imagen de una clula humana en mitosis, en la que los microtbulos se muestran en verde (formando el huso
mittico), los cromosomas en azul en el ecuador del huso y los cinetocoros en rojo.
El cinetocoro es una estructura proteica donde se anclan los microtbulos (MTs) del huso
mittico durante los procesos de divisin celular (meiosis y mitosis). Est localizado en una
zona especfica del cromosoma, el centrmero. En vertebrados y levaduras los cinetocoros
son estructuras discretas y nicas en cada cromosoma, pero existen organismos (como C.
elegans) que presentan cinetocoros difusos a lo largo de los brazos cromosmicos: son los
denominados cromosomas holocntricos.
Los cinetocoros inician, controlan y supervisan los llamativos movimientos de los
cromosomas durante la divisin celular. En cuanto a su estructura, los cinetocoros de las
clulas animales pueden subdividirse en dos regiones:
-el cinetocoro interno se organiza normalmente sobre secuencias de ADN altamente
repetido (el ADN satlite) y se ensambla en una forma especializada de cromatina que
persiste a travs del ciclo celular
-el cinetocoro externo es una estructura proteica con muchos componentes dinmicos
que se ensambla y funciona slo durante la divisin celular
Las funciones del cinetocoro incluyen el anclaje de los cromosomas a los MTs del huso
mittico, la verificacin de esos anclajes, la activacin del checkpoint de mitosis (un
mecanismo de control que retrasa la salida de mitosis si se detectan fallos) y la participacin
en la generacin de las fuerzas que propulsan los movimientos cromosmicos durante la
divisin celular.
Instituto Hernando Arias de Saavedra
BIOLOGA
63
Metafase
A medida que los microtbulos encuentran y se anclan a los cinetocoros durante la
prometafase, los centrmeros de los cromosomas se congregan en la "placa metafsica" o
"plano ecuatorial", una lnea imaginaria que es equidistante de los dos centrosomas que se
encuentran en los dos polos del huso. Este alineamiento equilibrado en la lnea media del
huso se debe a las fuerzas iguales y opuestas que se generan por los cinetocoros
hermanos. El nombre "metafase" proviene del griego que significa "despus."
Dado que una separacin cromosmica correcta requiere que cada cinetocoro est
asociado a un conjunto de microtbulos (que forman las fibras cinetocricas), los
cinetocoros que no estn anclados generan una seal para evitar la progresin prematura
hacia anafase antes de que todos los cromosomas estn correctamente anclados y
alineados en la placa metafsica. Esta seal activa el checkpoint de mitosis. Los
mecanismos que verifican que se cumplen los requisitos necesarios para pasar a la fase
siguiente del ciclo celular se denominan checkpoints o puntos de control. A travs del
ciclo celular, existen diferentes puntos de control. El checkpoint que asegura que la
segregacin cromosmica tiene lugar de forma correcta se denomina checkpoint de
mitosis, de anafase o tambin punto de control del ensamblaje del huso, abreviado SAC
por sus siglas en ingls (Spindle Assembly Checkpoint).
Metafase: Los cromosomas se encuentran alineados en la placa metafsica
Instituto Hernando Arias de Saavedra
BIOLOGA
64
Anafase
Cuando todos los cromosomas estn correctamente anclados a los microtbulos del huso y
alineados en la placa metafsica, la clula procede a entrar en anafase (del griego que
significa "arriba", "contra", "atrs" o "re-").
Entonces tienen lugar dos sucesos. Primero, las protenas que mantenan unidas ambas
cromtidas hermanas (las cohesinas
12
), son cortadas, lo que permite la separacin de las
cromtidas. Estas cromtidas hermanas, que ahora son cromosomas hermanos diferentes,
son separados por los microtbulos anclados a sus microtbulos al desensamblarse,
dirigindose hacia los centrosomas respectivos.
A continuacin, los microtbulos no asociados a cinetocoros se alargan, empujando a los
centrosomas (y al conjunto de cromosomas que tienen asociados) hacia los extremos
opuestos de la clula. Este movimiento parece estar generado por el rpido ensamblaje de
los microtbulos.
Estos dos estadios se denominan a veces anafase temprana (A) y anafase tarda (B). La
anafase temprana viene definida por la separacin de cromtidas hermanas, mientras que la
tarda por la elongacin de los microtbulos que produce la separacin de los centrosomas.
Al final de la anafase, la clula ha conseguido separar dos juegos idnticos de material
gentico en dos grupos definidos, cada uno alrededor de un centrosoma.
Anafase: los microtbulos anclados a cinetocoros se acortan y los dos juegos de cromosomas se aproximan a
cada uno de los centrosomas.
Telofase
La telofase (del griego , que significa "finales") es la reversin de los procesos que
tuvieron lugar durante profase y prometafase. Durante la telofase, los microtbulos no
12
La cohesina es un complejo de protenas involucrado en la separacin de las cromtidas, encargada de
mediar la unin de las cromtidas en la metafase. Es uno de los complejos proteicos responsables de mantener
la estructura de los cromosomas.
Instituto Hernando Arias de Saavedra
BIOLOGA
65
unidos a cinetocoros continan alargndose, estirando an ms la clula. Los cromosomas
hermanos se encuentran cada uno asociado a uno de los polos. La membrana nuclear se
reforma alrededor de ambos grupos cromosmicos, utilizando fragmentos de la membrana
nuclear de la clula original. Ambos juegos de cromosomas, ahora formando dos nuevos
ncleos, se descondensan de nuevo en cromatina. La cariocinesis ha terminado, pero la
divisin celular an no est completa.
Telofase: Los cromosomas decondensados estn rodeados por la membrana nuclear.
Instituto Hernando Arias de Saavedra
BIOLOGA
66
Consecuencias de la mitosis
Mediante el proceso mittico, el material gentico se divide en dos ncleos idnticos, con lo
que las dos clulas hijas que resultan si se produce la divisin del citoplasma (citocinesis)
sern genticamente idnticas. Por tanto, la mitosis es un proceso de divisin conservativo,
ya que el material gentico se mantiene de una generacin celular a la siguiente. La mayor
parte de la expresin gnica
13
se detiene durante la mitosis, pero mecanismos
epigenticos
14
funcionan durante esta fase, para "recordar" los genes que estaban activos
en mitosis y transmitirlos a las clulas hijas.
Errores en la mitosis
Aunque los errores en la mitosis son bastante poco frecuentes, este proceso puede fallar,
especialmente durante las primeras divisiones celulares en el cigoto
15
. Los errores mitticos
pueden ser especialmente peligrosos para el organismo, porque el descendiente futuro de la
clula madre defectuosa mantendr la misma anomala.
Un cromosoma puede no separarse durante la anafase. Este fenmeno se denomina "no-
disyuncin". Si esto ocurre, una clula hija recibir dos cromosomas hermanos y la otra se
quedar sin ninguno. Esto da lugar a que una clula tenga tres cromosomas que codifiquen
la misma cosa (dos hermanos y un homlogo), una condicin conocida como trisoma
16
, y la
13
La expresin gnica es el proceso por medio del cual todos los organismos procariotas y eucariotas
transforman la informacin codificada en los cidos nucleicos en las protenas necesarias para su desarrollo y
funcionamiento. En todos los organismos, inclusive los eucariotas, el contenido del ADN de todas sus clulas es
idntico. Esto quiere decir que contienen toda la informacin necesaria para la sntesis de todas las protenas.
Pero no todos los genes se expresan al mismo tiempo ni en todas las clulas.
14 La epigentica es el estudio de modificaciones en la expresin de genes que no se encuentra en la
secuencia del ADN y estas modificaciones son heredables. Una de las fuentes de mayores modificaciones de los
genes es por el factor ambiental y puede afectar a uno o varios genes con mltiples funciones. Por medio de la
regulacin epigentica se puede observar como es la adaptacin al medio ambiente dada por la plasticidad del
genoma el cual tiene como resultado la formacin de distintos fenotipos dependientes del medio ambiente al que
sea expuesto el organismo. Estas modificaciones que se dan presentan un alto grado de estabilidad y al ser
heredables esto permite que se puedan mantener es un linaje celular por muchas generaciones. Esto es
importante ya que cuando hay errores en las modificaciones se pueden generar enfermedades que perduren en
una familia por mucho tiempo.
15 En biologa, se denomina cigoto (zigoto, huevo) a la clula resultante de la unin del gameto masculino con
el femenino en la reproduccin sexual de los organismos (animales, plantas, hongos y algunos eucariotas
unicelulares). Su citoplasma y sus orgnulos son siempre de origen materno al proceder del vulo.
16 En gentica, una trisoma es la existencia de un cromosoma extra en un organismo diploide: en vez de un
par homlogo de cromosomas es un triplete (2n + 1 cromosomas). Los efectos de las trisomas van en paralelo a
los de las monosomas. La adicin de un cromosoma extra da lugar a individuos algo ms viables que en el
caso de la prdida de un cromosoma. La variacin de los cromosomas sexuales del tipo trisoma tiene un efecto
menos grave en el fenotipo que las variaciones autosmicas. En la especie humana, la adicin de un
cromosoma extra X o Y a una mujer o un varn da lugar a individuos viables que presentan diversos sndromes.
Instituto Hernando Arias de Saavedra
BIOLOGA
67
otra clula, que solamente tiene un cromosoma (el cromosoma homlogo), tendr
monosoma
17
. Estas clulas se consideran aneuploides, y la aneuploida
18
puede causar
inestabilidad gentica, un hecho frecuente en cncer.
Monosoma parcial en la especie humana
En la especie humana no se han documentado monosomas autosmicas ms all del
nacimiento. Los individuos con tales dotaciones cromosmicas, que no sobreviven al
desarrollo embrionario y fetal. Sin embrago, hay casos de supervivientes con monosomas
parciales, en donde se pierde solo parte de un cromosoma (deleciones segmentales).
Jerome Le Jeune en 1963, describi los sntomas clnicos del sndrome del Cri-du-Chat.
Este sndrome est asociado con la perdida de una pequea parte del brazo corto del
cromosoma 5.
La adicin de un autosoma grande a la dotacin diploide tiene graves efectos y normalmente es letal durante
su desarrollo.
17
En Gentica, la prdida de un cromosoma da lugar a un complemento cromosmico 2n - 1 y tal condicin se
denomina monosoma. La incapacidad para sobrevivir de los individuos monosmicos en muchas especies
animales es desconcertante, ya que al menos hay una copia de cada gen en el homlogo restante. Una
explicacin posible se refiere al desenmascaramiento de letales recesivos que son tolerados en los heterocigotos
que llevan los correspondientes alelos silvestres. Si un organismo heterocigoto para un solo gen letal recesivo
pierde el cromosoma homlogo que lleva el alelo normal, el cromosoma desapareado dar lugar a la muerte del
organismo. Otra explicacin posible es que la informacin gentica en el desarrollo temprano est regulada
cuidadosamente, de tal manera que se requiere un delicado equilibrio de los productos gnicos para asegurar un
desarrollo normal Ejemplo: maz, tabaco, hierba del asno y entre otras plantas. Tales plantas monosmicas son
casi siempre menos viables que las diploides de donde provienen.
18 En gentica, el trmino aneuploida hace referencia a cambios en el nmero de cromosomas, que pueden
dar lugar a enfermedades genticas. Un aneuploide es un individuo cuyo nmero de cromosomas difiere del tipo
salvaje en parte de su dotacin cromosmica, debido a un cromosoma extra o ausente, que siempre se asocia
con una deficiencia en el desarrollo fsico, mental o ambos. Generalmente, la dotacin cromosmica aneuploide
slo difiere de la salvaje en uno o pocos cromosomas. La aneuploida se puede observar frecuentemente en
clulas cancerosas. En los animales slo son viables las monosomas y las trisomas, ya que las nulisomas son
letales en individuos diploides.
Instituto Hernando Arias de Saavedra
BIOLOGA
68
Esquema del genoma tras una mutacin, en este caso una trisoma del cromosoma 21.
Trisomas
Las trisomas ms frecuentes en los seres
humanos son:
Sndrome de Down
Sndrome de Down, que es la aneuploida
ms viable, con un 0,15% de individuos en la
poblacin. Es una trisoma del cromosoma 21
(aunque puede producirse por translocacin),
que incluye retraso mental (CI de 20-50), cara
ancha y achatada, estatura pequea, ojos con
pliegue epicntico y lengua grande y arrugada.
Es la nica trisoma autosmica de la especie
humana de la que sobrevive un nmero
significativo de individuos ms all del ao
despus del nacimiento fue descubierta por
Langdon Down en 1866. El sndrome es
consecuencia de la trisoma del cromosoma 21,
un cromosoma del grupo G1, y se llama
sndrome de Down, o simplemente trisoma
del 21. Los enfermos con sndrome de Down
presentan estatura baja, cabeza redondeada,
frente alta y aplanada, y lengua y labios secos
y fisurados. Presentan epicanto, pliegue de piel
en la esquina interna de los ojos. Las palmas
de las manos muestran un nico pliegue
transversal, y las plantas de los pies presentan
un pliegue desde el taln hasta el primer
espacio interdigital (entre los dos primeros
dedos). En muchos casos padecen
cardiopatas congnitas y tienden a desarrollar
leucemia. El cociente de inteligencia (CI) vara
desde 20 hasta 60 (una inteligencia media
alcanza el valor 100), pero con procedimientos
educativos especficos y precoces, algunos
enfermos consiguen valores ms altos. La
incidencia global del sndrome de Down se
Instituto Hernando Arias de Saavedra
BIOLOGA
69
aproxima a uno de cada 700 nacimientos, pero
el riesgo vara con la edad de la madre. La
incidencia en madres de 25 aos es de 1 por
2000 nacidos vivos, mientras que en madres de
35 aos es de 1 por cada 200 nacimientos y de
1 por cada 40 en las mujeres mayores de 40
aos. Para detectar la anormalidad
cromosmica durante el periodo prenatal se
pueden emplear la amniocentesis y la biopsia
de vellosidades corinicas. Algunas
alteraciones sanguneas maternas pueden
sugerir la gestacin de un hijo con sndrome de
Down: niveles bajos de alfa-fetoprotena y
niveles anormales de estriol no conjugado y
gonadotrofina corinica humana.
La anomala cromosmica causante de la
mayora de los casos de sndrome de Down es
la trisoma del 21, presencia de tres copias de
este cromosoma. Por tanto, los pacientes
presentan 47 cromosomas en vez de 46 (cifra
normal del genoma humano) en todas sus
clulas. Esta anomala es consecuencia de la
fertilizacin de un vulo patolgico de 24
cromosomas por un espermatozoide normal de
23 cromosomas, aunque tambin a veces la
anomala es generada por el espermatozoide.
En una clula germinal, la pareja de
cromosomas 21 se mantiene unida y pasa a
uno slo de los dos vulos o espermatozoides
derivados de ella. En un tipo ms raro de
sndrome de Down, producido por
translocacin, parte del material gentico de
uno de los cromosomas 21 se queda adherido
al otro de los cromosomas 21. Algunos
enfermos presentan alteraciones
cromosmicas slo en algunas clulas de su
organismo, no en todas; en este caso se dice
que presentan un mosaicismo.
Sindrome de Patau. Klaus Patau y sus
colaboradores (1960) observaron un nio con
graves malformaciones del desarrollo y un
cariotipo de 47 cromosomas. El cromosoma
extra tena un tamao medio, uno de los
acrocntricos del grupo D, que ahora se
denomina cromosoma 13. La trisoma del 13,
que se denomina sndrome de Patau (47,
+13). Los nios se cree que son sordos, tienen
una caracterstica fisura labio-palatina y
polidactilia. Las autopsias han revelado
malformaciones congnitas en muchos
sistemas orgnicos, indicativas del desarrollo
anormal que se produce a las cinco o seis
semanas de gestacin. El promedio de
supervivencia de estos nios es de unos tres
meses. La edad promedio de los padres de
nios afectados es mayor que la de los padres
de los nios normales, pero no es tan alta
como la edad promedio materna en los casos
del sndrome de Down. Este caso slo ocurre
en 1 de cada 19.000 nacimientos.
Sndrome de Edwards. John H. Edwards y
sus compaeros (1960) publicaron el caso de
un nio trismico para un cromosoma del grupo
E, que ahora se sabe es el cromosoma 18.
Esta aberracin de la trisoma del 18 (47, +18)
tambin se denomina sndrome de Edwards.
El fenotipo de estos nios, ilustra que la
presencia de un autosoma extra produce
malformaciones congnitas y una menor
esperanza de vida. Estos nios son ms
pequeos de lo normal. Sus crneos estn
alargados segn el eje antero-posterior y sus
orejas son deformes y situadas ms bajo de lo
normal. El cuello ancho, dislocacin congnita
de la cadera y mentn deprimido. El promedio
de supervivencia es menos de cuatro meses.
La muerte se produce normalmente por
neumona o fallo cardaco. La edad media
materna es alta, los bebs con el sndrome de
Edwards son preponderantemente mujeres.
Sndrome de Klinefelter (XXY). Produce
individuos altos, con fsico ligeramente
feminizado, cociente intelectual algo reducido,
disposicin femenina del vello del pubis, atrofia
testicular y desarrollo mamario. Tenemos una
mezcla de ambos sexos (individuos
ginandromorfos).
Trisoma 9. Es una trisoma del cromosoma 9.
Sndrome de Warkany. Es una trisoma del
cromosoma 8.La trisoma 8 es una anomala
Instituto Hernando Arias de Saavedra
BIOLOGA
70
cromosmica que en la mayora de los casos
descritos corresponde a un mosaico. Sus
caractersticas clnicas varan desde dismorfias
discretas hasta malformaciones severas que,
por lo general, incluyen retardo mental -leve a
grave-, dismorfias faciales tpicas, alteraciones
esquelticas, pliegues palmares y plantares
profundos, anomalas renales y otras.
Trisoma del 16. Es la trisoma ms frecuente,
ya que se da en el 1% de las mujeres
embarazadas, pero totalmente inviable, dando
lugar a un aborto alrededor del tercer mes.
Sndrome del triple X. Presenta tres
cromosomas X.
Sndrome del XYY. Tambin existen
aneuploides somticos, que son individuos
constituidos por diferentes lneas celulares con
diferente nmero de cromosomas. Se
denominan quimeras y se producen por una
no-disyuncin en la mitosis. Al principio del
desarrollo puede originarse un individuo
mosaico, como los ginandromorfos a nivel
sexual. Son individuos con cromosomas de
ambos sexos, pudiendo existir individuos
X0/XYY o XX/XY.
La mitosis es un proceso traumtico. La clula pasa por cambios drsticos en su
estructura, algunos orgnulos se desintegran y se reconstruyen en cuestin de horas, y
los microtbulos tiran constantemente de los cromosomas. Por tanto, en ocasiones los
cromosomas pueden daarse. Un brazo del cromosoma se puede romper y perder un
fragmento, causando delecin
19
. El fragmento puede incorporarse incorrectamente a otro
cromosoma no homlogo, causando translocacin
20
. Se puede integrar de nuevo al
19
Una delecin, en gentica, es un tipo especial de anomala estructural cromosmica que consiste en la
prdida de un fragmento de ADN de un cromosoma. Esta prdida origina un desequilibrio, por lo que las
deleciones estn incluidas dentro de las reordenaciones estructurales desequilibradas. El portador de una
delecin es monosmico respecto a la informacin gnica del segmento correspondiente del homlogo
normal. Una delecin puede producirse en el extremo de un cromosoma (delecin terminal) o a lo largo de
uno de sus brazos (delecin intersticial). El origen de las deleciones puede ser una sencilla rotura
cromosmica y prdida del segmento acntrico. En ciertos casos, las deleciones son el resultado de un
entrecruzamiento desigual entre cromosomas homlogos o cromtidas hermanas mal alineados. Tambin se
pueden producir en la descendencia por segregacin anormal de una translocacin o una inversin
equilibradas de los progenitores.
20
En Gentica, una translocacin cromosmica es el desplazamiento de un segmento de un cromosoma a
un nuevo lugar en el genoma. El intercambio de segmento entre dos cromosomas no homlogos es una
translocacin reciproca. El modo ms fcil para que ocurra esto es que los brazos de dos cromosomas no
homlogos se aproximen de tal manera que se facilite el intercambio. Si el intercambio incluye segmentos
cromosmicos internos, se necesitan cuatro roturas, dos en cada cromosoma. La translocacin robertsoniana
o fusin cntrica, implica roturas en el extremo final de los brazos cortos de dos cromosomas acrocntricos
no homlogos. Los pequeos fragmentos acntricos se pierden y los segmentos cromosmicos grandes se
fusionan por sus regiones centromricas, dando lugar a un nuevo cromosoma grande, metacntrico o
submetacentrico. Existen diversos mecanismos mediante los cuales dos cromosomas acrocntricos dan lugar
a un cromosoma meta o submetacntrico (fusin) y el caso recproco (Fisin). Una translocacin como esta
explica los casos familiares en los que el sndrome de Down se hereda. Los estudios citogenticos de los
Instituto Hernando Arias de Saavedra
BIOLOGA
71
cromosoma original, pero en una orientacin inversa, causando inversin. O se puede
tratar errneamente como un cromosoma separado, causando duplicacin
cromosmica
21
.
Tipos de mutaciones cromosmicas
padres y de sus descendientes en estos casos raros explican la causa del sndrome de Down familiar. Los
anlisis revelan que uno de los padres tiene una translocacin 14/21 D/G. Es decir, un padre tiene la mayor
parte del cromosoma 21 del grupo G translocado al extremo del cromosoma 14 del grupo D. este individuo es
normal, aun cuando l o ella tengan 45 cromosomas.
21 En Gentica, una duplicacin cromosmica es la repeticin de un fragmento de cromosoma a
continuacin del fragmento original. Las duplicaciones surgen por error en la duplicacin del ADN, como
producto de una reorganizacin cromosmica de tipo estructural o relacionado con un proceso de
sobrecruzamiento defectuoso. Las duplicaciones no suelen ser deletreas, son una fuente de nuevo material
gentico y base para nuevos cambios evolutivos. Muchas de las familias gnicas con un origen evolutivo
comn, o las familias multignicas pueden tener su origen en las duplicaciones. Si el segmento afectado es
de gran tamao, se puede detectar en meiosis con los mismos criterios que en las deleciones (bivalente
heteromorfo o zona intersticial desapareada en el cromosoma con la duplicacin).
Instituto Hernando Arias de Saavedra
BIOLOGA
72
Translocacin recproca entre los cromosomas 4 y 20 en humanos.
Delecin de un cromosoma.
Una parte de estos errores pueden detectarse por alguno de los puntos de control
existentes a travs del ciclo celular, lo cual produce una parada en la progresin celular,
dando tiempo a los mecanismos reparadores a corregir el error. Si esto no ocurre, el
efecto de estas anormalidades genticas depender de la naturaleza especfica del error.
Puede variar de una anomala imperceptible, a carcinognesis o a la muerte del
organismo.
Instituto Hernando Arias de Saavedra
BIOLOGA
73
Endomitosis
La endomitosis es una variante de la mitosis sin divisin nuclear o celular, lo que da lugar
a clulas con muchas copias del mismo cromosoma en el mismo ncleo. Este proceso
tambin se denomina endoreduplicacin, y las clulas resultantes endoploides. Un
ejemplo de una clula que sufre endomitosis es el megacariocito.
La apoptosis es una forma de muerte celular, que est regulada genticamente.
La muerte celular programada es parte integral del desarrollo de los tejidos tanto de
plantas (viridiplantae) como de animales pluricelulares (metazoa). En animales, la forma
de muerte celular programada ms corriente es la "apoptosis". Cuando una clula muere
por apoptosis, empaqueta su contenido citoplasmtico, lo que evita que se produzca la
respuesta inflamatoria caracterstica de la muerte accidental o necrosis. En lugar de
hincharse o reventar y por lo tanto, derramar su contenido intracelular daino enzimtico,
hacia el espacio extracelular-, las clulas en proceso de apoptosis y sus ncleos se
encogen, y con frecuencia se fragmentan conformando vesculas pequeas que
contienen el material citoplasmtico. De esta manera, pueden ser eficientemente
englobadas va fagocitosis y, consecuentemente, sus componentes son reutilizados por
macrfagos o por clulas del tejido adyacente.
Ciclo celular y cncer
Instituto Hernando Arias de Saavedra
BIOLOGA
74
Cuando las clulas normales se lesionan o envejecen, mueren por apoptosis, pero las
clulas cancerosas la evitan.
Se cree que muchos tumores son el resultado de una multitud de pasos, de los que una
alteracin mutagnica no reparada del ADN podra ser el primer paso. Las alteraciones
resultantes hacen que las clulas inicien un proceso de proliferacin descontrolada e
invadan tejidos normales. El desarrollo de un tumor maligno requiere de muchas
transformaciones genticas. La alteracin gentica progresa, reduciendo cada vez ms la
capacidad de respuesta de las clulas al mecanismo normal regulador del ciclo.
8
Los genes que participan de la carcinognesis resultan de la transformacin de los genes
normalmente implicados en el control del ciclo celular, la reparacin de daos en el ADN
y la adherencia entre clulas vecinas. Para que la clula se transforme en neoplsica se
requieren, al menos, 2 mutaciones: una en un gen supresor de tumores y otra en un
protooncogn, que d lugar, entonces, a un oncogn.
Citocinesis
Esquema de citocinesis
Instituto Hernando Arias de Saavedra
BIOLOGA
75
Una clula a punto de completar la citocinesis. Le flecha seala un centrosoma.
La citocinesis es un proceso independiente, que se inicia simultneamente a la telofase.
Tcnicamente no es parte de la mitosis, sino un proceso aparte, necesario para
completar la divisin celular. En las clulas animales, se genera un surco de escisin
(cleavage furrow) que contiene un anillo contrctil de actina en el lugar donde estuvo la
placa metafsica, estrangulando el citoplasma y aislando as los dos nuevos ncleos en
dos clulas hijas. Tanto en clulas animales como en plantas, la divisin celular est
dirigida por vesculas derivadas del aparato de Golgi, que se mueven a lo largo de los
microtbulos hasta la zona ecuatorial de la clula. En plantas esta estructura forma una
placa celular en el centro del fragmoplasto y se desarrolla generando una pared celular
que separa los dos ncleos. El fragmoplasto es una estructura de microtbulos tpica de
plantas superiores, mientras que algunas algas utilizan un vector de microtbulos
denominado ficoplasto durante la citocinesis. Al final del proceso, cada clula hija tiene
una copia completa del genoma de la clula original. El final de la citocinesis marca el
final de la fase M.
Esquema resumen de las distintas fases de la divisin celular: profase, prometafase, metafase, anafase,
telofase y citocinesis.
REPASAMOS
La citocinesis o citodiresis es la separacin fsica del citoplasma en dos clulas hijas
durante la divisin celular. Se produce despus de la cariocinesis, y al final de la telofase,
en la divisin celular mittica. Su mecanismo es distinto en la clula animal (por
estrangulamiento) o vegetal (por tabicacin).
Instituto Hernando Arias de Saavedra
BIOLOGA
76
En clulas animales la formacin de un surco de divisin implica una expansin
de la membrana en esta zona y una contraccin progresiva causada por un anillo
perifrico contrctil de actina asociada a miosina. Este anillo producir la
separacin de las dos clulas hijas por estrangulacin del citoplasma.
Las clulas vegetales tienen un proceso diferente de divisin que consiste en la
acumulacin de vesculas procedentes del aparato de Golgi, que contienen
elementos de la pared celular, en la zona media de la clula. Las vesculas se
fusionan y entran en contacto con las paredes laterales de la clula. De esta
forma se origina el tabique o fragmoplasto que har posible la divisin celular.
Citocinesis animal
La citocinesis de la clula animal comienza poco despus de la separacin de cromtidas
hermanas en la anafase de la mitosis. A partir de miosinas II no musculares y de
filamentos de actina se forma meridionalmente un anillo contrctil en el cortex celular
(adyacente a la membrana celular). La miosina II utiliza la energa libre liberada cuando
se hidroliza el ATP para moverse a lo largo de los filamentos de actina, obligando a la
membrana celular a formar un surco de segmentacin. La hidrlisis continuada provoca
que el surco ingrese hasta que se forma una estructura llamada cuerpo medio y el
proceso de abscisin segmenta a este ltimo en dos. La abcisin depende de filamentos
de septina bajo el surco de segmentacin que conforman una base estructural para
asegurar que la citocinesis se completa. Despus de la citocinesis, microtbulos no
cinetocoros se reorganizan formando un nuevo citoesqueleto cuando el ciclo celular
vuelve a la interfase.
Posicin del anillo contrctil
La posicin del anillo Cytokinesis in Animal Cells", Cambridge University Press
(1996)</ref> lo que parece depender de la GTPase RhoA, que provoca varios efectores
cascada (como las protenas kinasa ROCK y citron) para promover la activacin de la
miosina en una regin particular del cortex celular.
El huso central
Junto con la formacin del anillo contrctil, se forma el "huso central", una estructura
tambin basada en microtbulos. Muchas especies requieren el huso central para cumplir
eficientemente la citocinesis.
Regulacin temporal de la citocinesis
La citocinesis est regulada temporalmente para asegurar que slo ocurre despus de la
separacin de las cromatidas hermanas durante divisiones celulares normales. Para
lograrlo, muchos componentes de la maquinaria citocintica estn regulados para
asegurar que son capaces de cumplir una funcin particular en un estadio particular del
ciclo celular.
Instituto Hernando Arias de Saavedra
BIOLOGA
77
Meiosis
En biologa, meiosis (del griego , disminucin) es una de las formas de
reproduccin celular. Es un proceso de divisin celular en el cual una clula diploide (2n)
experimenta dos divisiones sucesivas, con la capacidad de generar cuatro clulas
haploides (n). Este proceso se lleva a cabo en dos divisiones nucleares y citoplasmticas,
llamadas: primera y segunda divisin meitica o simplemente meiosis I y meiosis II.
Ambas comprenden profase, metafase, anafase y telofase.
Durante la meiosis I los miembros de cada par homlogo de cromosomas se emparejan
durante la profase, formando bivalentes. Durante esta fase se forma una estructura
proteica denominada complejo sinaptonmico, permitiendo que se produzca la
recombinacin entre ambos cromosomas homlogos. Posteriormente se produce una
gran condensacin cromosmica y los bivalentes se sitan en la placa ecuatorial durante
la primera metafase, dando lugar a la migracin de n cromosomas a cada uno de los
polos durante la primera anafase.
Esta divisin reduccional es la responsable del mantenimiento del nmero cromosmico
caracterstico de cada especie.
En la meiosis II, las cromtidas hermanas que forman cada cromosoma se separan y se
distribuyen entre los ncleos de las clulas hijas. Entre estas dos etapas sucesivas no
existe la etapa S (replicacin del ADN). La maduracin de las clulas hijas dar lugar a
los gametos.
Historia de la meiosis
La meiosis fue descubierta y descrita por primera vez en 1876 por el conocido bilogo
alemn Oscar Hertwig (1849-1922), estudiando los huevos del erizo de mar.
Instituto Hernando Arias de Saavedra
BIOLOGA
78
Fue descrita otra vez en 1883, en el nivel de cromosomas, por el zologo belga Edouard
Van Beneden (1846-1910) en los huevos de los gusanos parsitos Ascaris. En 1887
observ que en la primera divisin celular que llevaba a la formacin de un huevo, los
cromosomas no se dividan en dos longitudinalmente como en la divisin celular asexual,
sino que cada par de cromosomas se separaba para formar dos clulas, cada una de las
cuales presentaba tan solo la mitad del nmero usual de cromosomas. Posteriormente,
ambas clulas se dividan de nuevo segn el proceso asexual ordinario. Van Beneden
denomin a este proceso meiosis.
El significado de la meiosis para la reproduccin y la herencia, sin embargo, no se
describi hasta 1890, cuando el bilogo alemn August Weismann (1834-1914) observ
que eran necesarias dos divisiones celulares para transformar una clula diploide en
cuatro clulas haploides si deba mantenerse el nmero de cromosomas. En 1911 el
genetista estadounidense Thomas Hunt Morgan (1866-1945) observ el
sobrecruzamiento en la meiosis de la mosca de la fruta, proporcionando as la primera
interpretacin segura y verdadera sobre la meiosis.
Meiosis y ciclo vital
La reproduccin sexual se caracteriza por la fusin de dos clulas sexuales haploides
para formar un cigoto diploide, por lo que se deduce que, en un ciclo vital sexual, debe
ocurrir la meiosis antes de que se originen los gametos.
En los animales y en otros pocos organismos, la meiosis precede de manera inmediata a
la formacin de gametos. Las clulas somticas de un organismo individual se
multiplican por mitosis y son diploides; las nicas clulas haploides son los gametos.
Estos se forman cuando algunas clulas de la lnea germinal experimentan la meiosis. La
formacin de gametos recibe el nombre de gametognesis. La gametognesis
masculina, denominada espermatognesis, conduce a la formacin de cuatro
espermatozoides haploides por cada clula que entra en la meiosis.
En contraste, la gametognesis femenina, llamada ovognesis, genera un solo vulo por
cada clula que entra en la meiosis, mediante un proceso que asigna virtualmente todo el
citoplasma a uno solo de los dos ncleos en cada divisin meitica. Al final de la primera
divisin meitica se retiene un ncleo; el otro, llamado primer cuerpo polar, se excluye de
la clula y por ltimo degenera. De modo similar, al final de la segunda divisin un ncleo
se convierte en el segundo cuerpo polar y el otro ncleo sobrevive. De esta forma, un
ncleo haploide pasa a ser el receptor de la mayor parte del citoplasma y los nutrimentos
acumulados de la clula meitica original.
Sin embargo, aunque la meiosis se realiza en algn punto de los ciclos vitales sexuales,
no siempre precede directamente a la formacin de gametos. Muchos eucariontes
sencillos (incluso algunos hongos y algas) permanecen haploides (sus clulas se dividen
por mitosis) la mayor parte de su vida, y los individuos pueden ser unicelulares o
Instituto Hernando Arias de Saavedra
BIOLOGA
79
pluricelulares. En ellos, dos gametos haploides (producidos por mitosis) se fusionan para
formar un cigoto diploide, que experimenta la meiosis para volver al estado haploide.
Los ciclos vitales ms complejos se encuentran en vegetales y en algunas algas. Estos
ciclos vitales, que se caracterizan por alternancia de generaciones, consisten en una
etapa diploide multicelular, denominada generacin esporfita, y una etapa haploide
multicelular, a la que se llama generacin gametfita. Las clulas esporofitas diploides
experimentan la meiosis para formar esporas haploides, cada una de las cuales se divide
en forma mittica para producir un gametofito haploide multicelular. Los gametofitos
producen gametos por mitosis. Los gametos femeninos y masculinos (vulos y
espermatozoides) se fusionan entonces para formar un cigoto diploide, el cual se divide
de manera mittica para producir un esporofito diploide multicelular.
Proceso celular
Visin general de la meiosis. En la interfase se duplica el material gentico, y se produce el fenmeno de la
recombinacin (representado por cromosomas rojos y azules). En meiosis I los cromosomas homlogos se
reparten en dos clulas hijas. En meiosis II, al igual que en una mitosis, cada cromtida migra hacia un polo.
El resultado son 4 clulas hijas haploides (n).
Los pasos preparatorios que conducen a la meiosis son idnticos en patrn y nombre a la
interfase del ciclo mittico de la clula. La interfase se divide en tres fases:
Fase G1: caracterizada por el aumento de tamao de la clula debido a la
fabricacin acelerada de organelas, protenas y otras materias celulares.
Fase S: se replica el material gentico, es decir, el ADN se replica dando origen a
dos cadenas nuevas, unidas por el centrmero. Los cromosomas, que hasta el
momento tenan una sola cromtida, ahora tienen dos. Se replica el 98% del ADN,
el 2% restante queda sin replicar.
Instituto Hernando Arias de Saavedra
BIOLOGA
80
Fase G2: la clula contina aumentando su biomasa.
La interfase es seguida inmediatamente por la meiosis I y II. La meiosis I consiste en la
segregacin de cada uno de los cromosomas homlogos, dividiendo posteriormente la
clula diploide en dos clulas diploides pero con la mitad de cromosomas. La meiosis II
consiste en desemparejar cada una de las cromtidas del cromosoma, que se
segregarn una a cada polo, con lo que tras una divisin se producen cuatro clulas
haploides. Meiosis I y II estn divididas en profase, metafase, anafase y telofase,
similares en propsito a sus subfases anlogas en el ciclo mittico de la clula. Por lo
tanto, la meiosis abarca la interfase (G1, S, G2), la meiosis I (profase I, metafase I,
anafase I, telofase I), y la meiosis II (profase II, metafase II, anafase II, telofase II).
Meiosis I
En meiosis I, los cromosomas en una clula diploide se segregan nuevamente,
produciendo cuatro clulas hijas haploides. Este es el paso de la meiosis que genera
diversidad gentica.
Profase I
La Profase I de la primera divisin meitica es la etapa ms compleja del proceso y a su
vez se divide en 5 subetapas, que son:
Leptoteno
La primera etapa de Profase I es la etapa del leptoteno, durante la cual los cromosomas
individuales comienzan a condensar en filamentos largos dentro del ncleo. Es diploide
2n.
Cigoteno
Los cromosomas homlogos comienzan a acercarse hasta quedar apareados en toda su
longitud. Esto se conoce como sinapsis (unin) y el complejo resultante se conoce como
bivalente o ttrada (nombre que prefieren los citogenetistas), donde los cromosomas
homlogos (paterno y materno) se aparean, asocindose as cromtidas homlogas.
Producto de la sinapsis, se forma una estructura observable solo con el microscopio
electrnico, llamada complejo sinaptonmico, unas estructuras, generalmente esfricas,
aunque en algunas especies pueden ser alargadas.
La disposicin de los crommeros a lo largo del cromosoma parece estar determinado
genticamente. Tal es as que incluso se utiliza la disposicin de estos crommeros para
poder distinguir cada cromosoma durante la profase I meitica. Durante el zigoteno
concluye la replicacin del ADN
Instituto Hernando Arias de Saavedra
BIOLOGA
81
Paquiteno
Una vez que los cromosomas homlogos estn perfectamente apareados formando
estructuras que se denominan bivalentes se produce el fenmeno de entrecruzamiento
(crossing-over) en el cual las cromtidas homlogas no hermanas intercambian material
gentico. La recombinacin gentica resultante hace aumentar en gran medida la
variacin gentica entre la descendencia de progenitores que se reproducen por va
sexual.
Diploteno
Los cromosomas continan condensndose hasta que se pueden comenzar a observar
las dos cromtidas de cada cromosoma. Adems en este momento se pueden observar
los lugares del cromosoma donde se ha producido la recombinacin. Estas estructuras en
forma de X reciben el nombre quiasmas. Cada quiasma se origina en un sitio de
entrecruzamiento, lugar en el que anteriormente se rompieron dos cromtidas homlogas
que intercambiaron material gentico y se reunieron.
Diacinesis
Esta etapa apenas se distingue del diploteno. Podemos observar los cromosomas algo
ms condensados y los quiasmas. El final de la diacinesis y por tanto de la profase I
meitica viene marcado por la rotura de la membrana nuclear. Durante toda la profase I
continu la sntesis de ARN en el ncleo. Al final de la diacinesis cesa la sntesis de ARN
y desaparece el nuclolo.
Etapas de la Profase I
Leptoteno:
En esta fase, los cromosomas se hacen visibles, como hebras largas y finas.
Otro aspecto de la fase leptoteno es el desarrollo de pequeas reas de
engrosamiento a lo largo del cromosoma, llamadas crommeros, que le dan
la apariencia de un collar de perlas.
Intercambio de fragmentos entre cromtidas
homlogas por entrecruzamiento de cromosomas
homlogos.
Instituto Hernando Arias de Saavedra
BIOLOGA
82
Cigoteno:
Es un perodo de apareamiento activo en el que se hace evidente que la
dotacin cromosmica del meiocito corresponde de hecho a dos conjuntos
completos de cromosomas. As, pues, cada cromosoma tiene su pareja,
cada pareja se denomina par homlogo y los dos miembros de la misma se
llaman cromosomas homlogos.
Paquiteno:
Esta fase se caracteriza por la apariencia de los cromosomas como hebras
gruesas indicativas de una sinapsis completa. As, pues, el nmero de
unidades en el ncleo es igual al nmero n.
A menudo, los nuclolos son muy importantes en esta fase. Los
engrosamientos cromosmicos en forma de perlas, estn alineados de forma
precisa en las parejas homlogas, formando en cada una de ellas un patrn
distintivo
Diploteno:
Ocurre la duplicacin longitudinal de cada cromosoma homlogo, al ocurrir
este apareamiento las cromtidas homlogas parecen repelerse y separarse
ligeramente y pueden apreciarse unas estructuras llamadas quiasmas entre
las cromtidas. La aparicin de estos quiasmas nos hace visible el
entrecruzamiento ocurrido en esta fase.
Diacinesis:
Esta etapa no se diferencia sensiblemente del diploteno, salvo por una
mayor contraccin cromosmica. Los cromosomas de la interfase, en forma
de largos filamentos, se han convertido en unidades compactas mucho ms
manejables para los desplazamientos de la divisin meitica.
Anotaciones de la Profase I
La membrana nuclear desaparece. Un cinetocoro se forma por cada cromosoma, no uno
por cada cromtida, y los cromosomas adosados a fibras del huso comienzan a moverse.
Algunas veces las ttradas son visibles al microscopio. Las cromtidas hermanas
continan estrechamente alineadas en toda su longitud, pero los cromosomas homlogos
ya no lo estn y sus centrmeros y cinetocoros se encuentran separados.
Instituto Hernando Arias de Saavedra
BIOLOGA
83
Metafase I
Los cromosomas homlogos se alinean en el plano de ecuatorial. La orientacin es al
azar, con cada homologo paterno en un lado. Esto quiere decir que hay posibilidad de
que las clulas hijas reciban el homlogo del padre o de la madre por cada cromosoma.
Anafase I
Los quiasmas se separan. Los microtbulos del huso se acortan en la regin del
cinetocoro, con lo que se consigue remolcar los cromosomas homlogos a lados
opuestos de la clula, junto con la ayuda de protenas motoras. Ya que cada cromosoma
homlogo tiene solo un cinetocoro, se forma un juego haploide (n) en cada lado. En la
reparticin de cromosomas homlogos, para cada par, el cromosoma materno se dirige a
un polo y el paterno al contrario. Por tanto el nmero de cromosomas maternos y
paternos que haya a cada polo vara al azar en cada meiosis. Por ejemplo, para el caso
de una especie 2n = 4 puede ocurrir que un polo tenga dos cromosomas maternos y el
otro los dos paternos; o bien que cada polo tenga uno materno y otro paterno.
Telofase I
Cada clula hija ahora tiene la mitad del nmero de cromosomas pero cada cromosoma
consiste en un par de cromtidas. Los microtbulos que componen la red del huso
mittico desaparece, y una membrana nuclear nueva rodea cada sistema haploide. Los
cromosomas se desenrollan nuevamente dentro de la cromatina. Ocurre la citocinesis
(proceso paralelo en el que se separa la membrana celular en las clulas animales o la
formacin de esta en las clulas vegetales, finalizando con la creacin de dos clulas
hijas). Despus suele ocurrir la intercinesis, parecido a una segunda interfase, pero no
es una interfase verdadera, ya que no ocurre ninguna rplica del ADN. No es un proceso
universal, ya que si no ocurren las clulas pasan directamente a la metafase II.
Meiosis II
La meiosis II es similar a la mitosis. Las cromtidas de cada cromosoma ya no son
idnticas en razn de la recombinacin. La meiosis II separa las cromtidas produciendo
dos clulas hijas, cada una con 23 cromosomas (haploide), y cada cromosoma tiene
solamente una cromtida.
Instituto Hernando Arias de Saavedra
BIOLOGA
84
Profase II
Profase Temprana II
Comienza a desaparecer la envoltura nuclear y el nuclolo. Se hacen evidentes largos
cuerpos filamentosos de cromatina, y comienzan a condensarse como cromosomas
visibles.
Profase Tarda II
Los cromosomas continan acortndose y engrosndose. Se forma el huso entre los
centrolos, que se han desplazado a los polos de la clula.
Metafase II
Las fibras del huso se unen a los cinetocoros de los cromosomas. stos ltimos se
alinean a lo largo del plano ecuatorial de la clula. La primera y segunda metafase
pueden distinguirse con facilidad, en la metafase I las cromtidas se disponen en haces
de cuatro (ttrada) y en la metafase II lo hacen en grupos de dos (como en la metafase
mittica). Esto no es siempre tan evidente en las clulas vivas.
Anafase II
Las cromtidas se separan en sus centrmeros, y un juego de cromosomas se desplaza
hacia cada polo. Durante la Anafase II las cromtidas, unidas a fibras del huso en sus
Instituto Hernando Arias de Saavedra
BIOLOGA
85
cinetocros, se separan y se desplazan a polos opuestos, como lo hacen en la anafase
mittica. Como en la mitosis, cada cromtida se denomina ahora cromosoma.
Telofase II
En la telofase II hay un miembro de cada par homologo en cada polo. Cada uno es un
cromosoma no duplicado. Se reensamblan las envolturas nucleares, desaparece el huso
acromtico, los cromosomas se alargan en forma gradual para formar hilos de cromatina,
y ocurre la citocinesis. Los acontecimientos de la profase se invierten al formarse de
nuevo los nuclolos, y la divisin celular se completa cuando la citocinesis ha producidos
dos clulas hijas. Las dos divisiones sucesivas producen cuatro ncleos haploide, cada
uno con un cromosoma de cada tipo. Cada clula resultante haploide tiene una
combinacin de genes distinta. Esta variacin gentica tiene dos fuentes:
1 Durante la meiosis, los cromosomas maternos y paternos se barajan, de modo que
cada uno de cada par se distribuye al azar en los polos de la anafase I.
2 - se intercambian segmentos de ADN entre los homlogos paternos y maternos durante
el entrecruzamiento.
La Meiosis en imgenes
Profase I (temprana) Profase I (intermedia) Profase I (tarda)
En el cito primario los cromosomas se
ven como filamentos muy delgados
Los cromosomas homlogos se
aparean (sinapsis) y se hacen ms
cortos y gruesos
Cada cromosoma tiene dos
cromtidas hermanas unidas por
un centrmero. La membrana
nuclear empieza a desaparecer
Metafase I Anafase I Telofase I
Las ttradas se ordenan en el ecuador Los cromosomas homlogos se Se forman dos ncleos haploides.
Instituto Hernando Arias de Saavedra
BIOLOGA
86
del huso separan, dirigindose a los polos
opuestos. Los centrmeros no se
dividen
Cada cromosoma consta de dos
cromtidas adheridas a un
centrmero
Citos secundarios Profase II Metafase II
Durante la intercinesis no hay
duplicacin de material gentico
En los citos secundarios los
cromosomas se recondensan. La
membrana nuclear comienza a
desaparecer y se reconstituye el huso
acromtico
Los cromosomas se alinean en
el ecuador del huso como en la
mitosis
Anafase II Telofase II
Los centrmeros se dividen, separndose las
cromtidas hermanas. Los nuevos cromosomas
migran hacia los polos opuestos
Reconstruccin de los ncleos. Se completa la
citoquinesis formndose cuatro clulas haploides
que entran al periodo de interfase
Significado e importancia de la Meiosis
La meiosis no es un tipo de divisin celular diferente de la mitosis o una alternativa a
sta. La meiosis tiene objetivos diferentes.
Uno de estos objetivos es la reduccin cromosmica. Las clulas diploides se convierten
en haploides.
Otro de sus objetivos es establecer reestructuraciones en los cromosomas homlogos
mediante intercambios de material gentico. Por lo tanto, la meiosis no es una simple
divisin celular. La meiosis est directamente relacionada con la sexualidad y tiene un
profundo sentido para la supervivencia y evolucin de las especies.
A nivel gentico, la meiosis es una de las fuentes de variabilidad de la informacin.
Instituto Hernando Arias de Saavedra
BIOLOGA
87
Bsicamente, la meiosis es un mecanismo indispensable para asegurar la
constancia del nmero especfico de cromosomas en los organismos
sexuados.
Ya se ha visto que las dos divisiones meiticas reducen la cantidad de cromosomas del
nmero diploide (2n) (dos juegos de cromosomas) al haploide (n) (un juego de
cromosomas), lo que posibilita la unin de dos tipos diferentes de gametos para originar
un cigoto diploide (con los dos juegos de cromosomas).
Si la produccin de gametos se hiciera por mitosis, la fusin de ellos duplicara el nmero
cromosmico del cigoto. As, en la especie humana con 46 cromosomas por clula, la
unin del vulo y el espermatozoide dara lugar a un huevo con 92 cromosomas.
Al repetirse el mismo proceso, las generaciones sucesivas duplicaran indefinidamente la
cantidad de material cromosmico en cada clula, de manera que la prole siguiente
poseera 184 cromosomas, la subsiguiente 368 y, al llegar a la dcima generacin, los
individuos tendran sus clulas con 23.552 cromosomas en los ncleos. Esta
acumulacin continua de material cromosmico hara imposible la existencia de cualquier
clula.
Adems de garantizar la permanencia del nmero especfico de
cromosomas, la meiosis es muy importante porque provee la
continuidad del material hereditario de una generacin a la siguiente y,
a la vez, contribuye a crear variabilidad en la descendencia.
El "entrecruzamiento" de los cromosomas paternos y maternos durante
la profase I y la "combinacin al azar" de esos mismos cromosomas en
la metafase I, determinan la produccin de una gran variedad de
gametos por cada progenitor.
Como los gametos masculino y femenino tambin se unen al azar para
formar un cigoto, se puede afirmar que este proceso de fusin y la
meiosis que le precede, son importantes fuentes de variabilidad dentro
de las especies que presentan reproduccin sexual.
La variacin en la descendencia constituye la base de los cambios
evolutivos que ocurren con el tiempo. Los individuos que, por sus caractersticas
hereditarias, pueden adaptarse mejor a las condiciones ambientales tienen mayores
oportunidades de sobrevivir y dejar ms descendientes que los individuos con rasgos
hereditarios menos favorables.
Las clulas
haploides
resultantes de
la meiosis se
van a convertir
en las clulas
sexuales
reproductoras:
los gametos o
en clulas
asexuales
reproductoras:
las esporas.
Instituto Hernando Arias de Saavedra
BIOLOGA
88
Variabilidad gentica
El proceso de meiosis presenta una vital importancia en los ciclos vitales ya que hay una
reduccin del nmero de cromosomas a la mitad, es decir, de una clula diploide (ej: 46
cromosomas en el ser humano) se forman clulas haploides (23 cromosomas). Esta
reduccin a la mitad permite que en la fecundacin se mantenga el nmero de
cromosomas de la especie. Tambin hay una recombinacin de informacin gentica,
que es heredada del padre y la madre; el apareamiento de los homlogos y consecuente
crossing-over permite el intercambio de informacin gentica. Por lo tanto el nuevo
individuo hereda informacin gentica nica y nueva, y no un cromosoma ntegro de uno
de sus parientes. Otra caracterstica importante en la significacin de la meiosis para la
reproduccin sexual, es la segregacin al azar de cromosomas maternos y paternos. La
separacin de los cromosomas paternos y maternos recombinados, durante la anafase I
y II, se realiza completamente al azar, hecho que contribuye al aumento de la diversidad
gentica. En la anafase I, por cada par de homlogos existen dos posibilidades: un
cromosoma puede ir a un polo mittico o al otro.
El nmero de combinaciones posibles por tanto se calcula 2n donde n es el nmero de
pares de cromosomas homlogos (variaciones con repeticin de n elementos en grupos
de 2). En el ser humano, que tiene 23 pares de cromosomas homlogos, tiene la
posibilidad de recombinacin con 223 = 8.388.608 combinaciones, sin tener en cuenta las
mltiples combinaciones posibilitadas por la recombinacin en el crossing-over.
Anomalas cromosmicas
En la meiosis debe tener lugar una correcta separacin de las cromtidas hacia los polos
durante la anafase, lo que se conoce como disyuncin meitica; cuando esto no ocurre,
o hay un retraso en la primera o segunda divisin meiticas, conduce a problemas en la
configuracin de los cromosomas, alterndose el nmero correcto de estos, es decir,
dejan de ser mltiplos del nmero haploide original de la especie, lo que se conoce como
aneuploida. Entre los problemas en el material gentico encontramos:
Nulisoma en la que falta un par de cromosomas homlogos (2n-2 cromosomas)
Monosoma (2n-1 cromosomas)
Trisoma (2n+1 cromosomas)
En los animales slo son viables monosomas y trisomas. Los individuos nulismicos no
suelen manifestarse, puesto que es una condicin letal en diploides.
Instituto Hernando Arias de Saavedra
BIOLOGA
89
Trisoma de los cromosomas sexuales. Fuente la Web.
Los individuos con un slo cromosoma (XO) tiene el sndrome de Turner. Los XXY poseen sndrome de
Klinefelter. Los que lleven slo un cromosoma Y son inviables y los XXX se denominan super hembras. Las
alteraciones en el nmero de cromosomas que solo afectan a uno o dos cromosomas del complemento
cromosmico, se denominan ANEUPLOIDES.
Monosoma
Monosoma autosmica: produce la muerte en el tero.
Sndrome de Turner: solamente un cromosoma X presente. Los afectados son
hembras estriles, de estatura baja y un repliegue membranoso entre el cuello y
los hombros. Poseen el pecho con forma de escudo y pezones muy separados,
as como ovarios rudimentarios y manchas marrones en las piernas.
Trisoma
Sndrome de Down - Trisoma del cromosoma 21: es la aneuploida ms viable,
con un 0,15% de individuos en la poblacin. Incluye retraso mental (C.I de 20-50),
cara ancha y achatada, estatura baja, ojos con pliegue apicntico y lengua grande
y arrugada.
Sndrome de Patau - Trisoma del cromosoma 13. Se trata de la trisoma
menos frecuente. Se suele asociar con un problema meitico materno, ms que
paterno y, al igual que en el sndrome de Down, el riesgo aumenta con la edad de
Instituto Hernando Arias de Saavedra
BIOLOGA
90
la madre. Los afectados mueren poco tiempo despus de nacer, la mayora antes
de los 3 meses, como mucho llegan al ao. Se cree que entre el 80 y 90% de los
fetos con el sndrome no llegan a trmino.
Sndrome de Edwards - Trisoma del cromosoma 18. Es una enfermedad
infrecuente, que clnicamente se caracteriza por bajo peso al nacer, talla baja,
retraso mental y del desarrollo psicomotor (coordinacin de la actividad muscular
y mental), e hipertona (tono anormalmente elevado del msculo). Est
acompaada de diversas anomalas viscerales.
Sndrome de Klinefelter - Un cromosoma X adicional en varones (XXY).
Produce individuos altos, con fsico ligeramente feminizado, coeficiente intelectual
algo reducido, disposicin femenina del vello del pubis, atrofia testicular y
desarrollo mamario.
Sndrome del supermacho - Un cromosoma Y adicional en varones (XYY). No
presenta diferencias frente a los varones normales y de hecho se duda sobre el
uso del trmino sndrome para esta condicin.
Sndrome de la superhembra - Un cromosoma X adicional en mujeres. No
supone un riesgo aumentado de problemas de salud. Las mujeres con esta
condicin son altas, de bajo peso, con irregularidad en el periodo menstrual y rara
vez presentan debilidad mental.
Diferencia entre Mitosis y meiosis
Instituto Hernando Arias de Saavedra
BIOLOGA
91
ENTRE LA MITOSIS Y LA MEIOSIS
Cromosomas homlogos en mitosis (arriba) y meiosis (abajo).
(CUADRO RESUMEN)
MITOSIS MEIOSIS
A nivel gentico
Reparto exacto del material gentico. Segregacin al azar de los cromosomas
homlogos y entrecruzamiento como fuente de
variabilidad gentica.
A nivel celular
Como consecuencia de lo anterior se forman clulas
genticamente iguales.
Produce una reduccin del juego de cromosomas a
la mitad exacta de los cromosomas homlogos.
A nivel orgnico
Se da este tipo de divisin en los organismos
unicelulares para su reproduccin asexual y en
pluricelulares para su desarrollo, crecimiento y la
reparacin y regeneracin de tejidos y rganos.
Sirve para la formacin de las clulas
reproductoras sexuales: los gametos, o las clulas
reproductoras asexuales: las esporas.
Instituto Hernando Arias de Saavedra
BIOLOGA
92
Instituto Hernando Arias de Saavedra
BIOLOGA
93
Instituto Hernando Arias de Saavedra
BIOLOGA
94
Gametognesis
Gametognesis es la formacin de gametos por medio de la meiosis a partir de clulas
germinales. Mediante este proceso, el nmero de cromosomas que existe en las clulas
germinales se reduce de diploide a haploide, es decir, a la mitad del nmero de
cromosomas que contiene una clula normal de la especie de que se trate. En el caso de
los humanos si el proceso tiene como fin producir espermatozoides se le denomina
espermatognesis y se realiza en los testculos. En caso contrario, si el resultado, son
vulos se denomina ovognesis y se lleva a cabo en los ovarios.
Este proceso se realiza en dos divisiones cromosmicas y citoplasmticas, llamadas,
primera y segunda divisin meitica o simplemente Meiosis I y Meiosis II. Ambas
comprenden Profase, Prometafase, Metafase, Anafase, Telofase y Citocinesis. Durante la
meiosis I los miembros de cada par homlogo de cromosomas se unen primero y luego
se separan con el huso mittico y se distribuyen en diferentes polos de la clula. En la
Meiosis II, las cromtidas hermanas que forman cada cromosoma se separan y se
distribuyen en los ncleos de las nuevas clulas. Entre estas dos fases sucesivas no
existe la fase S (duplicacin del ADN).
La meiosis no es un proceso perfecto, a veces los errores en la meiosis son responsables
de las principales anomalas cromosmicas. La meiosis consigue mantener constante el
nmero de cromosomas de las clulas de la especie para mantener la informacin
gentica.
Gnadas
Tambin llamadas rganos sexuales primarios funcionan como glndulas mixtas en la
medida que se producen hormonas y gametos. Los rganos sexuales secundarios son
aquellas estructuras que maduran en la pubertad y que son esenciales en el cuidado y
transporte de gametos, son rasgos que se consideran de atraccin sexual.
Testculos: son dos estructuras ovaladas que se hallan suspendidas dentro del escroto
mediante cordones espermticos, son las que producen semen y lquido testicular; su
funcin endocrina es liberar hormonas masculinas como la testosterona, quienes
participaran en mantener los caracteres sexuales masculinos.
Ovarios: son dos rganos con forma de almendra, situados en los extremos de las
trompas de Falopio, los ovarios son formados aproximadamente cuando el feto hembra
tiene 3 meses y cuando la mujer entra a la pubertad los vulos se van desarrollando. Su
funcin endocrina es liberar hormonas como la progesterona y estrgeno, las cuales
intervendrn en el ciclo ovrico.
Instituto Hernando Arias de Saavedra
BIOLOGA
95
Funcin de las hormonas sexuales
Hombre, la testosterona es la principal hormona masculina, la sintetizan un grupo de
clulas llamadas clulas de Leyding, esta hormona promueve la espermatognesis o en
casos de abundancia la inhibe. El hipotlamo segrega el factor de liberacin por las
gonadotrofinas (GRF), el cual estimula la adenohipfisis y este a su vez estimule a la
Hipfisis para que libere la hormona luteinizante (LH) y la hormona folculo estimulante
(FSH).
Mujer, la hipfisis que produce la hormona FSH y la LH al igual que en el hombre, la cual
tiene accin sobre los ovarios, estimulando a un ovocito primario para completar la
meiosis I y formar un ovocito secundario. Al mismo tiempo la FSH produce estrgeno,
que produce el engrosamiento del endometrio. Al trmino de la FHS se produce la LH,
que produce la ovulacin. Las clulas germinales estn en ntimo contacto con el crvix y
el musculo sartorio el cual inerva e irriga a la misma vez al hueso humero.
Ovognesis
Proceso de formacin de gametos femeninos, que se localizan en los ovarios. Las
ovogonias se ubican en los folculos del ovario, crecen y tienen modificaciones; estos
llevan a la primera divisin meitica que da como resultado un ovocito primario (que
contiene la mayor parte del citoplasma) y un primer corpsculo polar. Las 2 clulas
resultantes efectan meiosis II, del ovocito secundario se forman una clula grande (que
tiene la mayor parte del citoplasma) y un segundo corpsculo polar, estos se desintegran
rpidamente, mientras que la clula grande se desarrolla convirtindose en los gametos
femeninas llamadas ovulo. Al ovulo lo rodean una capa de diferentes clulas, a esa capa
se le llama folculo de Graaf.
La ovogenesis cuenta con diversas fases las cuales son:
-Proliferacin: durante el desarrollo embrionario, las clulas germinales de los ovarios
sufren mitosis para originar a las ovogonias.
-Crecimiento: en la pubertad crecen para originar los ovocitos de 1er orden.
-Maduraciones ovocito del primer orden sufre meiosis
La ovogenesis comienza antes del nacimiento y se completa durante la vida
reproductiva de la mujer.
Espermatognesis
La espermatognesis es el mecanismo encargado de la produccin de
espermatozoides; es la gametognesis en el hombre. Este proceso se desarrolla en los
testculos, aunque la maduracin final de los espermatozoides se produce en el
Instituto Hernando Arias de Saavedra
BIOLOGA
96
epiddimo. La espermatognesis tiene una duracin aproximada de 64 a 75 das en la
especie humana, y consta de 3 fases o etapas:
- fase proliferativa,
-meiosis o espermatocitognesis, y
-espermiognesis o espermiohistognesis.
Diferencias entre Espermatognesis y Ovognesis
Espermatognesis
Se realiza en los testculos
Ocurre a partir de la espermatogonia
Cada espermatogonia da origen a cuatro espermatozoides
En la meiosis el material se divide equitativamente
Los espermatozoides se producen durante toda su vida
Se produce en el hombre
De un espermatocito I, se forman 4 espermios funcionales.
Ovognesis
Se realiza en los ovarios
Ocurre a partir de la ovogonia
Cada ovogonia da origen a un ovocito II el cual slo en el caso de ser fecundado
pasar a llamarse vulo y a 2 cuerpos polares I y a un cuerpo polar II (slo en
caso de fecundacin).
En meiosis I no se divide el citoplasma por igual, quedando una clula hija
(ovocito II) con casi todo el citoplasma
La mujer nace con un nmero determinado de Folculos, aproximadamente
400.000
Se produce en la mujer
De un ovocito I, se forma un vulo funcional.
Semejanzas entre Espermatognesis y Ovognesis
Ambos son sub-procesos de la gametognesis
Los 2 producen gametos
En ambos se produce la meiosis y mitosis
Los 2 son procesos de la reproduccin sexual en mamferos
Instituto Hernando Arias de Saavedra
BIOLOGA
97
Ambos procesos se producen dentro de las gnadas
Los 2 inician sus fases a partir de la mitosis
Comparacin entre vulos y espermatozoides
vulo (Ovocito II)
Ms grande que el espermatozoide
Tiene vitelo (reserva nutritiva)
No tiene movimiento
Sirve slo 1 de cada clula germinal
Se produce en el ovario
Espermatozoide
Pequeo en comparacin al ovocito II
No tiene reservas nutritivas
Se mueve por medio de su flagelo
Sirven 4 de cada clula germinal
Se produce en el testculo
Aspectos Histricos
El comienzo de las investigaciones en el desarrollo de las clulas reproductivas
masculinas coincide casi exactamente con la llegada de la teora celular, que
naturalmente conllevo al descubrimiento de la naturaleza celular de los espermatozoa;
la palabra aparece en 1827 por primera vez, sin embargo no es del todo claro quin fue el
primero en pronunciar la idea de que los espermatozoides provenan de clulas
testiculares. Koelliker (1841), quien colecto la primera evidencia notable acerca de este
tema, mantuvo que fue Rudolph Wagner, puesto que el examin los fluidos frescos
provenientes de los tbulos testiculares en mamferos, y los observ bajo el microscopio
evidenciando grnulos peculiares o esfrulas de formas y tamaos muy variables.
Wagner tambin observ Samenthierchen, o animlculos de esperma, espermatozoa,
sintindose seguro de que los varios tipos de esfrulas vistos con anterioridad eran
estados anteriores al espermatozoa. Su trabajo, en el que la palabra clula no ocurre ni
una sola vez, es caracterstico del tipo de concepto de formacin que precedi
inmediatamente a la teora celular2. Aos ms tarde, Albert Koelliker presenta un tratado
tras varios aos de investigacin, en el que ensea por primera vez los aspectos
fundamentales concernientes a la espermatogenesis:
Instituto Hernando Arias de Saavedra
BIOLOGA
98
1. En el semen de todos los animales, con algunas excepciones, se encuentran partculas
motiles Thelie, espermatozoa.
2. Los espermatozoa son la parte esencial del semen
3. Los espermatozoa se desarrollan individualmente en paquetes a partir de clulas que
se han formado en momentos de madurez sexual o de actividad testicular a travs de
procesos anlogos al desarrollo celular, pero significativamente diferentes del desarrollo
cigtico de los animales.
4. Las formas de los espermatozoa son bastante limitadas en variedad. Usualmente son
similares dentro de gneros, y frecuentemente tambin dentro de clases y familias. Cada
animal parece poseer solo un tipo de espermatozoa, con solo algunas excepciones.
Bajo estas conclusiones y un par mas, Koelliker deja claro que la visin del espermatozoa
como condicin primaria de un animal en desarrollo, sostenida por Leeuwenhoek y
muchos otros, deba ser abandonada2. A partir de ese momento, las investigaciones
acerca de la espermatognesis comenzaron a avanzar con mucha ms rapidez.
Finalmente, el descubrimiento ms importante en este campo y el de los tejidos
espermatognicos, fue la demostracin de las clulas de soporte (clulas de Sertoli), en
los tbulos seminferos de humanos hecha por el mismo hombre de cuyo nombre se
deriva el nombre de estas clulas2. Las corrientes de investigacin actuales estn
predominantemente orientadas a la resolucin de problemas humanos en el sentido ms
amplio de la palabra, y es llevada a cabo por especialistas en los campos mdicos y
paramdicos actuales, aun as, en el presente, es de suma importancia mantener la
significancia biolgica de este tema de manera firme en mente.
Anexo:
Espermatognesis
Los espermatozoides son clulas haploides que tienen la mitad de los cromosomas que
una clula somtica, son mviles y son muy diferenciadas. La reduccin en ellas se
produce mediante una divisin celular peculiar, la meiosis en el cul una clula diploide
(2n), experimentar dos divisiones celulares sucesivas sin un paso de duplicacin del
ADN entre dichas divisiones, con la capacidad de generar cuatro clulas haploides (n).
En este proceso es necesario pasar de unas clulas diploides, inmviles e
indiferenciadas a otras haploides, mviles y muy diferenciadas.
Un importante hecho a resaltar, es que mientras las divisiones reduccionales de la
meiosis se conservan en cada reino eukariota, la regulacin de la meiosis en mamferos
difiere dramticamente entre machos y hembras; entre estas diferencias, se encuentran:
- meiosis iniciada continuamente a partir de las poblaciones de clulas madre
correspondientes.
Instituto Hernando Arias de Saavedra
BIOLOGA
99
- 4 gametos producidos por ciclo mientras que en la ovognesis es solo 1 (un ovulo
funcional y 4 cuerpos polares).
- Meiosis completada en das o incluso semanas.
- La meiosis y diferenciacin procede continuamente sin arrestamiento del ciclo celular.
- La diferenciacin de gametos ocurre bajo un precursor haploide, luego de que la
meiosis termina.
- Los cromosomas sexuales son excluidos de recombinacin y transcripcin durante la
primera profase meitica.
La espermatognesis, en la especie humana, se produce en ondas a todo lo largo de los
tbulos seminferos, por lo que zonas adyacentes del mismo tbulo muestran
espermatocitognesis y espermiognesis en diversas fases. As, el proceso comienza
cuando las clulas indiferenciadas de los tbulos seminferos1 de los testculos se
multiplican.
Estas clulas germinales dan lugar a clulas madre de los espermatogonios, a partir de
las cuales surgen las clulas que se diferenciaran a espermatocitos primarios tras la
mitosis correspondiente de las clulas de tipo A. Las espermatogonias de tipo A,
entonces, a travs de repetidos ciclos de mitosis producen nuevas espermatogonias y
mantienen as la reserva celular. Estas clulas se caracterizan por poseer un gran ncleo
redondo u oval de cromatina condensada, en el que pueden encontrarse el ncleo
perifrico y una vacuola nuclear.
La produccin de espermatogonias de tipo A marca el inicio de la espermatognesis,
donde todas estas clulas son diploides.
Existen varios tipos de espermatogonias tipo A segn el aspecto de sus ncleos
celulares:
- Espermatogonias oscuras Ad, las cuales solo se dividen cuando tiene lugar una
reduccin drstica de espermatogonias, y adems se cree que estas clulas representan
las clulas madre del sistema y que su divisin mittica produce ms clulas de tipo Ad y
algunas de tipo Ap.
- Espermatogonias plidas Ap, las cuales dan lugar a ms espermatogonias del mismo
tipo o a espermatogonias de tipo B. Las primeras, dan lugar a clulas hijas unidas entre s
por puentes citoplasmticos, mientras que las segundas se originan por maduracin de
las primeras.
Las espermatogonias de tipo B se dividen por mitosis produciendo mas clulas de su
tipo; estas clulas, maduraran por grupos produciendo espermatocitos primarios y
eventualmente espermatozoides; tambin cabe resaltar, que las espermatogonias de este
tipo, se caracterizan por carecer de una vacuola nuclear. No se conoce aun que causa
que este tipo de espermatogonias siga el camino hacia la diferenciacin celular, antes
Instituto Hernando Arias de Saavedra
BIOLOGA
100
que el de auto-renovacin; tampoco es conocido que estimula a estas clulas para entrar
en divisin meiotica en vez de divisin mittica.
Cuando el individuo alcanza la madurez sexual, el proceso de espermatogenesis
regulado probablemente pos la sntesis de la protena BMP8B a partir de las propias
espermatogonias comienza cuando BMP8B alcanza niveles crticos de concentracion; las
espermatogonias de tipo B aumentan de tamao y se transforman en espermatocitos de
primer orden que migran al compartimiento adluminal del tbulo seminifero antes de
comenzar la primera divisin meitica mientras siguen siendo diploides: El ADN de los
espermatocitos primarios se replica justo despus de su formacin constituyendo un
estado 4n, y marcando el final de lo que se conoce como espermatocitogenesis.
Estas clulas, se reconocen fcilmente por sus abundantes citoplasmas y sus grandes
ncleos, que contienen grumos gruesos o finas hebras de cromatina1; adems, tambin
estn unidas por un puente citoplasmtico conspicuo como se mencion con anterioridad
que regula el proceso de divisin de forma que todas las clulas involucradas reciban la
seal de hacer meiosis a la vez mediante diferentes iones o molculas; dichas uniones
citoplasmticas solo se rompern una vez los espermatozoides sean liberados en la luz
del tbulo seminfero. As, es en estas clulas donde sucede la meiosis por primera vez:
La meiosis I dar lugar a dos espermatocitos de segundo orden; estos espermatocitos
secundarios, son entonces ms pequeos, y progresan con rapidez a la segunda divisin
meiotica, por lo que rara vez son visibles.
Tras la meiosis II resultarn cuatro espermtidas (gracias a la meiosis, de una clula
diploide surgen cuatro clulas haploides (gametos)); los gametos as producidos siguen el
proceso de maduracin conocido como espermiogenesis, donde en dicha fase las
espermtidas se convierten en espermatozoides; para ello, se reduce el citoplasma, el
ncleo de la clula se alarga y queda en la cabeza del espermatozoide generando la
forma puntiaguda caracterstica de este tipo de clulas. As, los espermatozoides
presentan tres zonas bien diferenciadas: la cabeza, el cuello y la cola. La primera es la de
mayor tamao, contiene los cromosomas de la herencia y lleva en su parte anterior un
pequeo saliente o acrosoma cuya misin es perforar las envolturas del vulo. En el
cuello se localiza el centrosoma y las mitocondrias, y el flagelo, que se origina a partir de
los centriolos, es el filamento que se encarga de generar la motilidad que le permite al
espermatozoide "nadar" hasta el vulo para [fecundacin|fecundarlo]]. El examen de los
distintos cortes de los tbulos de un testculo normal demuestra que alrededor de la mitad
de las clulas espermatogenicas se encuentra en estadio de espermatide tarda. Las
clulas de Sertoli, representan un componente de gran importancia en la regulacin de la
espermatognesis y espermatognesis. Durante el proceso de desarrollo, estas clulas
cuyos ncleos suelen encontrarse hacia la membrana basal del tbulo seminfero,
sostienen a las serie espermatognica. El ncleo de las clulas tiene una forma triangular
u oval tpica, con un ncleo prominente y cromatina dispersa.
Instituto Hernando Arias de Saavedra
BIOLOGA
101
La capa basal de las clulas germinales se adosa a la membrana basal, rodeada por una
lmina propia en la que existen varias capas de miofibroblastos y fibroblastos fusiformes.
Se forman uniones intimas tanto entre las mismas clulas, y estas con las clulas
germinales en desarrollo. Se sabe que para la produccin y maduracin de las clulas
espermatognicas es esencial que existan concentraciones elevadas de hormonas
andrognicas secretadas por las clulas de Leydig del mesotelio reticular. Las clulas de
Sertoli secretan una protena captadora de andrgenos que transporta la testosterona y la
dihidrotestosterona hacia la luz del tbulo seminfero. Dichas hormonas son tambin
necesarias para la funcin del epitelio de la rete testis y del epiddimo; la produccin de
esta protena de unin parece depender de una gonadotropina hipofisaria, la hormona
estimulante del folculo (FSH). Un ciclo espermatognico tiene una duracin aproximada
de 2 meses. Las espermatogonias estn en mitosis durante 16 das, dando lugar a los
espermatocitos primarios. Estos tardan 24 das en completar la primera meiosis y dar
lugar a los espermatocitos secundarios, los cuales tardarn horas en convertirse en
espermtides. Las espermtides tienen que diferenciarse, como se ha comentado
anteriormente, tardando otros 24 das para ese proceso. Si sumamos todo obtenemos
unos 64 das desde el paso de espermatogonias hasta espermatozoides.
Al realizarse la fecundacin, estos espermatozoides antes de salir pasan por el epiddimo
del testculo, donde se realiza la espermiohistognesis, y obtienen la acrosoma, un estilo
de casco en el espermatozoide hecho de enzimas, y una glicolema (capa), que la protege
del pH de la vagina. Esta capa (glicolema), se pierde en la diferenciacin natural, que
desaparece antes de llegar al vulo para lograr entrar en l con la fuerza del acrosoma.
Recordemos adems, que el espermatozoide est formado por una zona intermedia
donde se alojan numerosas mitocondrias que garantizan el aporte energtico, tambin
estn formados por un flagelo constituido por un filamento axial rodeado por una vaina
fibrosa, que permite la movilidad.
Periodo embrionario
Una vez las clulas germinales llegan a la cresta germinal del embrin masculino, se
incorporan a los cordones sexuales, donde se mantendrn hasta la madures, y perforaran
a fin de formar un orificio pasante que corresponder a los tbulos seminferos, y el
epitelio de dichos tbulos se diferenciara en clulas de Sertoli3. La formacin de
espermatozoides comienza alrededor del da 24 del desarrollo embrionario en la capa
endodrmica del saco vitelino. Aqu se producen unas 100 clulas germinales que migran
hacia los esbozos de los rganos genitales. Alrededor de la cuarta semana ya se
acumulan alrededor de 4000 de estas clulas, y el gen SRY determina que formen los
testculos para poder producir espermatozoides, aunque este proceso no empezar hasta
la pubertad. Hasta entonces, las clulas germinales se dividen por dos.
Instituto Hernando Arias de Saavedra
BIOLOGA
102
Accin Hormonal
Esta regulacin se produce por retroalimentacin negativa, desde el hipotlamo, el que
acta en la hipfisis, y finalmente en el testculo. Las hormonas responsables son:
Testosterona: responsable de las caractersticas sexuales masculinas, es
secretada en el testculo por las clulas de Leydig o instersticiales. Esta hormona
tambin es secretada por la mdula de la corteza suprarrenal pero en menores
cantidades en comparacin al testculo, tambin se secreta en las mujeres pero
en mnimas cantidades.
FSH u hormona folculo estimulante: secretada por la hipfisis, acta sobre las
clulas de Srtoli o nodrizas para que stas acten sobre los espermios en
desarrollo.
LH u hormona luteinizante: secretada por la hipfisis, acta sobre las clulas de
Leydig o instersticiales para que secreten testosterona.
Inhibina: secretada las clulas de Srtoli o nodrizas, acta sobre la hipfisis
inhibiendo la secrecin de FSH y con ello deteniendo la espermatogenesis.
Eficiencia
El hecho de que durante el eyaculado se produzca una elevada prdida de
espermatozoides, junto con un nmero bastante elevado de espermatozoides
malformados, convierte este mecanismo en un proceso redundante e ineficaz: de toda la
capacidad espermatognica potencial del testculo, tan slo un 25% consigue evitar la
prdida por apoptosis o degeneracin. A su vez, la mayora son malformados, motivo por
el cual nicamente un 12% de las clulas iniciales son potencialmente.
Ovognesis
La ovognesis es el proceso de formacin y diferenciacin de los gametos femeninos u
vulos en los animales, incluido el ser humano. La ovognesis, al igual que la
espermatognesis, se basa en el proceso de la meiosis, que produce, mediante dos
divisiones sucesivas, cuatro clulas con un genotipo recombinado y la mitad de ADN.
En los seres humanos, y en otros mamferos es ms o menos semejante, las clulas
germinales diploides generadas por mitosis, llamadas ovogonias (u oogonias), se
localizan en los folculos del ovario, crecen y sufren una diferenciacin para transformarse
en ovocitos primarios (u oocitos), donde se pone en marcha la primera divisin
meitica, dando origen una clula voluminosa u ovocito secundario que contiene la
mayor parte del citoplasma original y otra clula pequea o primer cuerpo polar (primer
corpsculo polar).
Instituto Hernando Arias de Saavedra
BIOLOGA
103
Estas dos clulas efectan la segunda divisin meitica; del ovocito secundario se forman
otras dos clulas: una grande, que contiene la mayor parte del citoplasma original, y otra
pequea o segundo cuerpo polar. Los cuerpos polares se desintegran rpidamente,
mientras que la otra clula se desarrolla para convertirse en un vulo maduro, haploide.
En los seres humanos el feto femenino empieza a formar ovogonias, pero se detiene el
proceso de meiosis en la etapa de ovocito primario, especficamente en profase I, en la
subfase conocida como diplotena. Este perodo se mantiene suspendido hasta que, a
partir de la pubertad y por efectos hormonales, se desprende un ovocito en cada ciclo
menstrual, se concluye entonces la primera divisin meitica y se inicia la segunda. sta
a su vez se interrumpe (en la metafase II), y no se completa hasta la fecundacin si es
que sta ocurre, cuando se forman el vulo y los tres corpsculos polares.
La reserva ovrica en mujeres, definida como el nmero de clulas germinales
(ovogonias durante el desarrollo y ovocitos tras el nacimiento) presenta el mximo a los 6
meses del comienzo del desarrollo embrionario, tras el cual desciende abruptamente
justo antes del nacimiento y contina decreciendo exponencialmente hasta la
menopausia. La cantidad y calidad de los ovocitos se puede relacionar con el posible
resultado reproductivo en mujeres cuando se realizan estudios de fertilidad, puesto que
en caso de presentar una reserva ovrica disminuida se obtendrn pocos ovocitos tras
tcnicas de induccin de ovulacin y, por tanto, poca probabilidad de embarazo positivo
Instituto Hernando Arias de Saavedra
BIOLOGA
104
Gentica
ADN, base de la herencia gentica.
La gentica (del trmino "Gen", que proviene de la palabra griega y significa
"descendencia") es una disciplina de la biologa, es la ciencia de la herencia y la variacin
en los seres vivos. El hecho de que los seres vivos heredan caracteres de sus padres ha
sido utilizado desde tiempos prehistricos para mejorar cultivos y animales mediante la
cra selectiva. Sin embargo, la ciencia moderna de la gentica, que aspira a comprender
el proceso de la herencia, slo empez con el trabajo de Gregor Mendel a mediados del
siglo XIX. Aunque no conoca la base fsica de la herencia, Mendel observ que los
organismos heredan caracteres de manera diferenciada estas unidades bsicas de la
herencia son actualmente denominadas genes.
Los genes corresponden a regiones del ADN, una molcula compuesta de una cadena de
cuatro tipos diferentes de nucletidos la secuencia de estos nucletidos es la
informacin gentica que heredan los organismos. El ADN existe naturalmente en forma
bicatenaria, es decir, en dos cadenas en que los nucletidos de una cadena
complementan los de la otra. Cada cadena puede hacer de plantilla para la creacin de
una nueva cadena complementaria este es el mtodo fsico para la creacin de copias
de genes que pueden ser heredados.
La secuencia de nucletidos de un gen es traducida por las clulas para producir una
cadena de aminocidos, creando protenas el orden de los aminocidos en una
protena corresponde con el orden de los nucletidos del gen. Esto recibe el nombre de
cdigo gentico. Los aminocidos de una protena determinan cmo se pliega en una
forma tridimensional, esta estructura es la responsable, a su vez, del funcionamiento de
Instituto Hernando Arias de Saavedra
BIOLOGA
105
la protena. Las protenas ejecutan casi todas las funciones que las clulas necesitan
para vivir. Un cambio en el ADN de un gen puede cambiar los aminocidos de una
protena, cambiando su forma y la funcin, lo que puede tener un efecto drstico sobre la
clula y el organismo en conjunto. Otros dos factores que pueden variar la forma de la
protena son el pH y la temperatura.
Aunque la gentica juega un papel significativo en la apariencia y el comportamiento de
los organismos, es la combinacin de la gentica con las experiencias del organismo la
que determina el resultado final. Por ejemplo, mientras que los genes juegan un papel en
la determinacin de la altura de una persona, la nutricin y la salud de esta persona
durante la infancia tambin juegan un papel importante.
El genoma es la totalidad de la informacin gentica que posee un organismo en
particular. Por lo general, al hablar de genoma en los seres eucariticos nos referimos
slo al ADN contenido en el ncleo, organizado en cromosomas. Pero no debemos
olvidar que tambin la mitocondria contiene genes (vase genoma mitocondrial). El
trmino fue acuado en 1920 por Hans Winkler, profesor de Botnica en la Universidad
de Hamburgo, Alemania, como un acrnimo de las palabras gene y chromosoma.
El trmino diploide indica que un organismo tiene dos copias del genoma en sus clulas,
debido a la presencia de pares de cromosomas homlogos.
El genoma no analiza la diversidad gentica o el polimorfismo de los genes de una
especie. Por ejemplo, en el genoma humano la secuencia en principio podra ser
determinada con slo la mitad del ADN de una clula de un individuo. Para conocer una
variacin particular o en enfermedades se requiere la comparacin entre individuos.
Subdivisiones de la gentica
La gentica se subdivide en varias ramas, como:
Clsica o mendeliana: Se preocupa del estudio de los cromosomas y los genes y
de cmo se heredan de generacin en generacin.
Cuantitativa, que analiza el impacto de mltiples genes sobre el fenotipo, muy
especialmente cuando estos tienen efectos de pequea escala.
Molecular: Estudia el ADN, su composicin y la manera en que se duplica.
Asimismo, estudia la funcin de los genes desde el punto de vista molecular.
de Poblaciones y evolutiva: Se preocupa del comportamiento de los genes en
una poblacin y de cmo esto determina la evolucin de los organismos.
del desarrollo: Se preocupa de cmo los genes controlan el desarrollo de los
organismos.
Instituto Hernando Arias de Saavedra
BIOLOGA
106
Ingeniera gentica
La ingeniera gentica es la especialidad que utiliza tecnologa de la manipulacin y
trasferencia del ADN de unos organismos a otros, permitiendo controlar algunas de sus
propiedades genticas. Mediante la ingeniera gentica se pueden potenciar y eliminar
cualidades de organismos en el laboratorio. Por ejemplo, se pueden corregir defectos
genticos (terapia gnica), fabricar antibiticos en las glndulas mamarias de vacas de
granja o clonar animales como la oveja Dolly. Algunas de las formas de controlar esto es
mediante transfeccin (lisar clulas y usar material gentico libre), conjugacin
(plsmidos) y transduccin (uso de fagos o virus), entre otras formas. Adems se puede
ver la manera de regular esta expresin gentica en los organismos.
Cuando los cientficos comprendieron la estructura de los genes y cmo la informacin
que portaban se traduca en funciones o caractersticas, comenzaron a buscar la forma
de aislarlos, analizarlos, modificarlos y hasta de transferirlos de un organismo a otro para
conferirle una nueva caracterstica. Justamente, de eso se trata la ingeniera gentica, un
conjunto de metodologas que permite transferir genes de un organismo a otro. Como
consecuencia, la ingeniera gentica sirve para clonar fragmentos de ADN y para
expresar genes (producir las protenas para las cuales estos genes codifican) en
organismos diferentes al de origen. As, es posible no slo obtener las protenas
recombinantes de inters sino tambin mejorar cultivos y animales. Hasta el momento se
ha utilizado la ingeniera gentica para producir, por ejemplo:
Vacunas, como la de la hepatitis B
Frmacos, como la insulina y la hormona del crecimiento humano
Enzimas para disolver manchas, como las que se usan en los detergentes en
polvo
Enzimas para la industria alimenticia, como las empleadas en la elaboracin del
queso y en la obtencin de jugos de fruta.
Plantas resistentes a enfermedades y herbicidas.
El desarrollo de la ingeniera gentica (tambin llamada metodologa del ADN
recombinante) fue posible gracias al descubrimiento de las enzimas de restriccin y de
los plsmidos. Las enzimas de restriccin reconocen secuencias determinadas en el
ADN. De esta manera, conociendo la secuencia de un fragmento de ADN es posible
aislarlo del genoma original para insertarlo en otra molcula de ADN. Hay muchas
enzimas de restriccin obtenidas a partir de bacterias y que sirven como herramientas
para la ingeniera gentica. Las enzimas de restriccin reconocen secuencias de 4, 6 o
ms bases y cortan generando extremos romos o extremos cohesivos. Estos extremos,
generados en diferentes molculas de ADN, pueden sellarse con la enzima ADN ligasa y
generar as una molcula de ADN nueva, denominada recombinante.
Instituto Hernando Arias de Saavedra
BIOLOGA
107
Los plsmidos son molculas de ADN circulares, originalmente aisladas de bacterias y
que pueden extraerse de las mismas e incorporarse a otras, a travs del proceso de
transformacin. Los plsmidos fueron modificados por los investigadores para ser
empleados como vectores. As, el gen de inters puede insertarse en el plsmido-vector
e incorporarse a una nueva clula. Para seleccionar las clulas (bacterias o clulas
animales o vegetales) que recibieron el plsmido, ste lleva, adems del gen de inters
(por ej., el gen de la insulina humana), un gen marcador de seleccin (por. ej., de
resistencia a un antibitico), que le otorga a la clula que lo lleva la capacidad de
sobrevivir en un medio de cultivo selectivo (medio con antibitico, en este ejemplo). Las
clulas que sobreviven se dividen y generan colonias, formadas por bacterias idnticas.
Estas bacterias se denominan recombinantes o genticamente modificadas. El plsmido
recombinante puede aislarse de estas colonias y transferirse a otras clulas.
Por esta metodologa es posible introducir genes de inters en todo tipo de clulas,
empleando los vectores y las tcnicas propias de cada sistema. Podemos entonces
generalizar los pasos de la ingeniera gentica de la siguiente manera:
1. Identificar un carcter deseable en el organismo de origen.
2. Encontrar el gen responsable del carcter deseado (gen de inters).
3. Combinar dicho gen con otros elementos necesarios (vector) para que
ste sea funcional en el organismo receptor.
4. Transferir el gen de inters, previamente introducido en el vector
adecuado, al organismo receptor.
5. Crecer y reproducir el organismo receptor, ahora modificado
genticamente.
Por ejemplo, para el caso de la transferencia de un gen insecticida de una bacteria al
maz:
1. Identificar la caracterstica resistencia a insectos en el organismo de origen,
la bacteria del suelo Bacillus thuringiensis.
2. Encontrar al gen que lleva las instrucciones para esta caracterstica.
3. Combinar este gen con otros elementos genticos para que sea funcional
ahora en una planta (ligarlo a un vector).
4. Transferir este gen a clulas de maz (organismo receptor).
5. Identificar las clulas de maz que recibieron el gen (clulas transformadas) y
regenerar, a partir de estas clulas, una planta adulta.
Instituto Hernando Arias de Saavedra
BIOLOGA
108
LECTURA
Qu es la Ingeniera Gentica?
De los genes a la ingeniera gentica
Cuando los cientficos comprendieron la estructura de los genes y cmo la
informacin que portaban se traduca en funciones o caractersticas, comenzaron a
buscar la forma de aislarlos, analizarlos, modificarlos y hasta de transferirlos de un
organismo a otro para conferirle una nueva caracterstica. Justamente, de eso se trata
la ingeniera gentica, que se podra definir como un conjunto de metodologas que
permite transferir genes de un organismo a otro y expresarlos (producir las protenas
para las cuales estos genes codifican) en organismos diferentes al de origen. El ADN
que combina fragmentos de organismos diferentes se denomina ADN recombinante.
En consecuencia, las tcnicas que emplea la ingeniera gentica se denominan
tcnicas de ADN recombinante. As, es posible no slo obtener protenas
recombinantes de inters sino tambin mejorar cultivos y animales. Los organismos
que reciben un gen que les aporta una nueva caracterstica se denominan
organismos genticamente modificados (OGM) o transgnicos. A su vez, la ingeniera
gentica es lo que caracteriza a la biotecnologa moderna que implementa estas
tcnicas en la produccin de bienes y servicios tiles para el ser humano, el ambiente
y la industria.
Etapas para la obtencin de un organismo transgnico
La siguiente tabla resume los pasos bsicos de la ingeniera gentica empleados para
transformar un organismo, y se ejemplifica con un caso concreto:
Metodologa Caso: obtencin de maz Bt que produce una
protena recombinante que le confiere resistencia a
determinados insectos
1.Identificar un carcter
deseable en el organismo de
origen
1. Identificar el carcter resistencia a insectos en
el organismo de origen, la bacteria del suelo
Bacillus thuringiensis (Bt)
2. Encontrar el gen
responsable del carcter
deseado (gen de inters),
aislarlo y caracterizarlo.
2. Encontrar al gen que lleva las instrucciones para
esta caracterstica, aislarlo y caracterizarlo.
3.Combinar dicho gen con
otros elementos necesarios
(vector) para que ste sea
funcional en el organismo
receptor
3. Combinar este gen con otros elementos
genticos para que sea funcional en una planta:
especialmente una secuencia promotora (y ligarlo a
un vector adecuado para transformar plantas)
Instituto Hernando Arias de Saavedra
BIOLOGA
109
4. Transferir el gen de inters,
previamente introducido en el
vector adecuado, al organismo
receptor.
4. Transferir este gen a clulas de maz (organismo
receptor).
5. Crecer y reproducir el
organismo receptor, ahora
modificado genticamente.
5. Identificar las clulas de maz que recibieron el
gen (clulas transformadas) y regenerar, a partir de
estas clulas, una planta adulta resistente a
insectos.
Tcnicas de Ingeniera Gentica o del ADN Recombinante
La obtencin de un organismo transgnico mediante tcnicas de ingeniera gentica
implica la participacin de un organismo que dona el gen de inters y un organismo
receptor del gen que expresar la nueva caracterstica deseada. Por ejemplo, para el
caso particular de la produccin de una variedad de maz que resista el ataque de
insectos, el organismo dador es la bacteria del suelo denominada Bacillus
thuringiensis (Bt) de la cual se extrae el gen que determina la sntesis de la protena
insecticida, y el organismo receptor del gen es la planta de maz. Las etapas y
tcnicas involucradas en este proceso seran:
1. Corroborar que existe un gen que codifica para la caracterstica de inters.
Cuando se encuentra una caracterstica en un organismo que resulta interesante para
transferir a otro organismo debe verificarse que es producto de un gen. Se identifica
el gen de inters por medio de cruzamientos a partir de una caracterstica que se
expresa, y se verifican las proporciones mendelianas. Si la caracterstica se atribuye a
una protena, que es producto directo de un gen, ser ms sencillo transferir esa
caracterstica a un organismo que no la tiene.
2. Clonar el gen de inters. Clonar un gen significa tenerlo puro en el tubo de
ensayos, o mejor an, dentro de un vector (una molcula mayor de ADN que permite
guardar fragmentos de ADN en forma estable y prctica por ms tiempo). La tarea de
clonar un gen involucra varias tcnicas: i) Extraccin de ADN; ii) Bsqueda de un gen
entre la mezcla de genes del ADN; iii) Secuenciacin; iv) Construccin del vector
recombinante. El ADN de inters se inserta en plsmidos-vectores que son
molculas de ADN lineales o circulares en las cuales se puede guardar (clonar) un
fragmento de ADN. Los ms usados son los plsmidos de origen bacteriano.
Instituto Hernando Arias de Saavedra
BIOLOGA
110
Los plsmidos pueden extraerse de las bacterias e incorporarse a otras, a travs del
proceso de transformacin. Los plsmidos fueron modificados por los investigadores
para ser empleados como vectores (vehculos). As, el gen de inters puede
insertarse en el plsmido-vector e incorporarse a una nueva clula
El desarrollo de estas tcnicas fue posible en gran medida por el descubrimiento de
las enzimas de restriccin. Las enzimas de restriccin reconocen secuencias
determinadas en el ADN. De esta manera, conociendo la secuencia de un fragmento
de ADN, es posible aislarlo del genoma original para insertarlo en otra molcula de
ADN. Hay muchas enzimas de restriccin obtenidas a partir de bacterias y que sirven
como herramientas para la ingeniera gentica
Instituto Hernando Arias de Saavedra
BIOLOGA
111
Las enzimas de restriccin reconocen secuencias de 4, 6 o ms bases y cortan
generando extremos romos o extremos cohesivos. Estos extremos, generados en
diferentes molculas de ADN, pueden sellarse con la enzima ADN ligasa y generar
as una molcula de ADN nueva, denominada recombinante.
Para tener gran cantidad y fcil disponibilidad del ADN de inters, el vector se inserta
dentro de bacterias (E. coli), las cuales crecen fcil y rpidamente. O sea, la bacteria
se utiliza como multiplicadora del vector, y por ende del inserto de inters. Esta es
una etapa de amplificacin del ADN para poder tener gran cantidad para
secuenciarlo, caracterizarlo, y luego poder hacer con l ADN recombinante.
En placas de Petri se cultivan las bacterias. Se originan colonias de bacterias iguales
(clones). Las bacterias transformadas, que incorporaron el plsmido y el gen de
inters, se seleccionan por medio de antibiticos y por reacciones con indicadores de
color.
. 3. Caracterizar el gen de inters. A partir de conocer la secuencia del gen se
puede, mediante bioinformtica, comparar esta secuencia con las de genes ya
conocidos para determinar a qu gen se parece, y se le asigna una posible funcin.
Una vez predicha la funcin del gen clonado por medio de anlisis informtico, se
debe proceder a confirmar la funcin real in vivo, o sea corroborar que en un sistema
biolgico funciona acorde a lo que se prev. Para ello se suele transferir el gen a un
organismo modelo, en el cual se pueda expresar el gen y medir su funcin. En el
ejemplo del maz, el gen Bt se puede transferir primero a las especies modelo
Arabidopsis thaliana y Nicotiana tabacum
4. Modificar el gen de inters. Si as se desea se puede agregar, deletar o mutar
secuencias dentro de la regin codificante, y agregar secuencias (promotor,
terminador, intrones) para que se pueda expresar en el sistema de inters. Por
ejemplo: si se clona un gen Bt de una bacteria para luego ponerlo en maz, se debe
agregar un promotor que funcione bien en plantas, es decir, que permita que las
clulas vegetales expresen la protena Bt. El promotor es una regin fundamental del
Instituto Hernando Arias de Saavedra
BIOLOGA
112
gen ya que determina cundo y dnde se expresar el gen.
EPGRAFE: Inserto preparado para ser transferido.
Fuente: http://www.agronort.com/informacion/ abcbiotec/abcbio6.html
5. Transformacin de un organismo con el gen de inters. Una vez hecha la
construccin gentica con el gen y promotor deseado, se elige el mtodo de
transformacin ms indicado para el organismo que se desea hacer transgnico.
6. Caracterizacin del OGM. Una vez obtenido el OGM, se lo analiza desde el punto
de vista molecular y biolgico. Para el anlisis molecular se debe demostrar, entre
otras cosas, si tiene una (o ms) copias del transgn, y cmo y en qu tejidos se
expresa el gen. Para analizar en qu tejido, momento y cantidad se expresa el gen se
analiza la presencia del ARN mensajero y de la protena recombinante codificados
por el transgn. Para la caracterizacin biolgica, el OGM se analiza desde el punto
de vista del objetivo (en este ejemplo, si el maz resulta efectivamente resistente a los
insectos) y desde el punto de vista que sea necesario acorde al OGM en cuestin. Si
ser utilizado como alimento y se lo cultivar a campo, entonces se deber hacer el
anlisis de riesgo alimentario y ambiental.
Hasta el momento se ha utilizado la ingeniera gentica para producir, entre otras
aplicaciones:
-Vacunas, por ejemplo contra la hepatitis B
-Frmacos, como la insulina y la hormona del crecimiento humano,
tanto en clulas transformadas y crecidas in vitro como en bacterias recombinantes y
animales transgnicos.
-Enzimas para disolver manchas, como las que se usan en los
detergentes en polvo, mayormente por medio de microorganismos recombinantes
(transgnicos) que crecen en biorreactores.
-Enzimas para la industria alimenticia, como las empleadas en la
Instituto Hernando Arias de Saavedra
BIOLOGA
113
elaboracin del queso y en la obtencin de jugos de fruta, entre otras.
-Plantas resistentes a enfermedades, entre otras caractersticas.
Cronologa de descubrimientos notables de la gentica
Ao Acontecimiento
1865 Se publica el trabajo de Gregor Mendel
1900
Los botnicos Hugo de Vries, Carl Correns y Eric Von Tschermak redescubren el trabajo de
Gregor Mendel
1903 Se descubre la implicacin de los cromosomas en la herencia
1905
El bilogo britnico William Bateson acua el trmino "Genetics" en una carta a Adam
Sedgwick
1910 Thomas Hunt Morgan demuestra que los genes residen en los cromosomas
1913 Alfred Sturtevant crea el primer mapa gentico de un cromosoma
1918
Ronald Fisher publica On the correlation between relatives on the supposition of Mendelian
inheritance la sntesis moderna comienza.
1923 Los mapas genticos demuestran la disposicin lineal de los genes en los cromosomas
1928
Se denomina mutacin a cualquier cambio en la secuencia nucleotdica de un gen, sea esta
evidente o no en el fenotipo
1928
Fred Griffith descubre una molcula hereditaria transmisible entre bacterias (vase
Experimento de Griffith)
1931 El entrecruzamiento es la causa de la recombinacin
1941
Edward Lawrie Tatum y George Wells Beadle demuestran que los genes codifican
protenas; vase el dogma central de la Gentica
1944
Oswald Theodore Avery, Colin McLeod y Maclyn McCarty demuestran que el ADN es el
material gentico (denominado entonces principio transformante)
1950
Erwin Chargaff demuestra que las proporciones de cada nucletido siguen algunas reglas
(por ejemplo, que la cantidad de adenina, A, tiende a ser igual a la cantidad de timina, T).
Barbara McClintock descubre los transposones en el maz
Instituto Hernando Arias de Saavedra
BIOLOGA
114
1952
El experimento de Hershey y Chase demuestra que la informacin gentica de los fagos
reside en el ADN
1953 James D. Watson y Francis Crick determinan que la estructura del ADN es una doble hlice
1956
Jo Hin Tjio y Albert Levan establecen que, en la especie humana, el nmero de
cromosomas es 46
1958
El experimento de Meselson y Stahl demuestra que la replicacin del ADN es
semiconservativa
1961 El cdigo gentico est organizado en tripletes
1964
Howard Temin demuestra, empleando virus de ARN, excepciones al dogma central de
Watson
1970
Se descubren las enzimas de restriccin en la bacteria Haemophilius influenzae, lo que
permite a los cientficos manipular el ADN
1977
Fred Sanger, Walter Gilbert, y Allan Maxam secuencian ADN por primera vez trabajando
independientemente. El laboratorio de Sanger completa la secuencia del genoma del
bacterifago -X174
1983
Kary Banks Mullis descubre la reaccin en cadena de la polimerasa, que posibilita la
amplificacin del ADN
1989
Francis Collins y Lap-Chee Tsui secuencian un gen humano por primera vez. El gen
codifica la protena CFTR, cuyo defecto causa fibrosis qustica
1990
Se funda el Proyecto Genoma Humano por parte del Departamento de Energa y los
Institutos de la Salud de los Estados Unidos
1995
El genoma de Haemophilus influenzae es el primer genoma secuenciado de un organismo
de vida libre
1996
Se da a conocer por primera vez la secuencia completa de un eucariota, la levadura
Saccharomyces cerevisiae
1998
Se da a conocer por primera vez la secuencia completa de un eucariota pluricelular, el
nematodo Caenorhabditis elegans
2001
El Proyecto Genoma Humano y Celera Genomics presentan el primer borrador de la
secuencia del genoma humano
2003
(14 de abril) Se completa con xito el Proyecto Genoma Humano con el 99% del genoma
secuenciado con una precisin del 99,99%
1
Instituto Hernando Arias de Saavedra
BIOLOGA
115
Herencia biolgica
La herencia biolgica es el proceso por el cual la prole de una clula u organismo
adquiere o est predispuesta a adquirir, las caractersticas de sus progenitores. El estudio
de la herencia biolgica es lo que se conoce como gentica, que incluye:
Herencia gentica, resultado de la replicacin del ADN y de la divisin celular.
Herencia epigentica resultado de, entre otras cosas, modificaciones en el ADN
como el "silencio" o no expresin de un gen.
Herencia gentica
La herencia gentica es la transmisin a travs del material gentico contenido en el
ncleo celular, de las caractersticas anatmicas, etc. de un ser vivo a sus descendientes.
El ser vivo resultante tendr caractersticas de uno o de los dos padres.
La herencia consiste en la transmisin a su descendencia de los caracteres de los
ascendentes. El conjunto de todos los caracteres transmisibles, que vienen fijados en los
genes, recibe el nombre de genotipo y su manifestacin exterior en el aspecto del
individuo el de fenotipo. Se llama idiotipo al conjunto de posibilidades de manifestar un
carcter que presenta un individuo.
Para que los genes se transmitan a los descendientes es necesaria una reproduccin
idntica que d lugar a una rplica de cada uno de ellos; este fenmeno tiene un lugar en
la meiosis.
Las variaciones que se producen en el genotipo de un individuo de una determinada
especie se denominan variaciones genotpicas. Estas variaciones genotpicas surgen
por cambios o mutaciones (espontneas o inducidas por agentes mutagnicos) que
pueden ocurrir en el ADN.
Las mutaciones que se producen en los genes de las clulas sexuales pueden
transmitirse de una generacin a otra. Las variaciones genotpicas entre los individuos de
una misma especie tienen como consecuencia la existencia de fenotipos diferentes.
Algunas mutaciones producen enfermedades, tales como la fenilcetonuria, galactosemia,
anemia falciforme, sndrome de Down, sndrome de Turner, entre otras. Hasta el
momento no se ha podido curar una enfermedad gentica, pero para algunas patologas
se est investigando esta posibilidad mediante la terapia gnica.
Lo esencial de la herencia queda establecido en la denominada teora cromosmica de
la herencia, tambin conocida como teora cromosmica de Sutton y Boveri:
1. Los genes estn situados en los cromosomas.
Instituto Hernando Arias de Saavedra
BIOLOGA
116
2. Los genes estn dispuestos linealmente en los cromosomas.
3. La recombinacin de los genes se corresponde con el intercambio de segmentos
cromosmicos. (Crossing over)
La transferencia gentica horizontal es factor de confusin potencial cuando se infiere un
rbol filogentico basado en la secuencia de un gen. Por ejemplo, dadas dos bacterias
lejanamente relacionadas que han intercambiado un gen, un rbol filogentico que
incluya a ambas especies mostrara que estn estrechamente relacionadas puesto que el
gen es el mismo, incluso si muchos de otros genes tuvieran una divergencia substancial.
Por este motivo, a veces es ideal usar otras informaciones para inferir filogenias ms
robustas, como la presencia o ausencia de genes o su ordenacin, o, ms
frecuentemente, incluir el abanico de genes ms amplio posible.
LECTURA
Artculo
En los aos 80 el cientfico Alec Jeffreys buscando cmo se heredaba la fibrosis qustica,
encontr la huella gentica que nos hace nicos. Pero Qu es la herencia gentica?
Sabemos por qu somos como somos? Controlaremos qu le legamos a nuestros hijos?
Tres 14, el programa de ciencia para toda la familia de TVE responder este domingo
estos interrogantes. Explicar por qu somos como somos y qu es lo que heredan los
hijos de los padres. A las 20,30 horas, en la 2 de TVE.
Una hermosa actriz le propuso al escritor irlands George Bernard tener un hijo. Tendra la belleza
de ella y la inteligencia de l, asegur la actriz. Shaw se neg. Saba que el experimento poda
salir al revs. El resultado gentico de un apareamiento es muy difcil de predecir. Pero an as
nos preguntamos por qu somos como somos. Queremos saber por qu tenemos los ojos del
abuelo o si determinada enfermedad se hereda. Y la respuesta a nuestras preguntas est en los
genes. Cromosomas, aminocidos y protenas son conceptos que a veces se nos mezclan en la
cabeza, pero que estn detrs de lo que nos hace tan iguales y tan diferentes. "Tres 14" ha
hablado con el bilogo Marcos Isamat y su familia, con la ginecloga Carmina Comas y con el
onclogo Manel Esteller. A ellos les preguntamos qu es la herencia? Sabemos por qu somos
como somos? Controlaremos qu le legamos a nuestros hijos?
El reportaje muestra una antigua fbrica de pan convertida en hogar, donde se ha presenta un
libro lleno de preguntas y respuestas. Su ttulo es Por qu mi hijo se parece a su abuela? y trata
de resolver las dudas que a todos nos asaltan al pensar en nuestros hijos. Quizs es este el nico
libro de divulgacin cientfica que se ha presentado en una casa particular, por la noche y con
nios en pijama firmando libros. Quizs es este el nico libro de ciencia familiar que se ha editado,
el menos en estos tiempos tan especializados y tecnificados. Lo cierto es que en pocas casas
convive un cientfico con una periodista. A travs de su charla descubrimos por qu somos como
somos.
Crticas a la definicin de herencia como herencia gentica
Instituto Hernando Arias de Saavedra
BIOLOGA
117
La Teora de los sistemas de desarrollo (DST) se opone a la definicin de herencia
como transmisin de genes y aplica el concepto a cualquier recurso que se encuentre en
generaciones sucesivas y que contribuya a explicar por qu cada generacin se parece a
la que le precede. Estos recursos incluyen factores celulares y factores externos como la
gravedad o la luz solar. La DST utiliza, por tanto, el concepto de herencia para explicar la
estabilidad de la forma biolgica de una generacin a otra.
Historia de la herencia biolgica
El primero en estudiar la herencia biolgica fue Gregor Mendel; mediante sus estudios
con guisantes y otros vegetales pudo enunciar tres leyes con las cuales explicaba el
modo de transmisin de ciertas caractersticas de los progenitores a la descendencia, y
que hoy da an se siguen teniendo en cuenta:
Primera ley, o Principio de la uniformidad: Cuando se cruzan dos individuos
de raza pura, los hbridos resultantes son todos iguales."
Segunda ley, o Principio de la segregacin: Un organismo individual posee dos
alelos que codifican un rasgo y que estos alelos se separan en proporciones
iguales cuando se forman gametos.
Tercera ley, o Principio de la combinacin independiente: Los genes que
codifican diferentes caractersticas se distribuyen de forma independiente cuando
se forman los gametos.
Herencia y cromosomas
A las caractersticas a las que se refera Mendel las denomin factores hereditarios.
Diversos estudios realizados a lo largo del siglo XX determinaron que stos estaban
localizados dentro del ncleo de las clulas, en concreto en unas partculas llamadas
cromosomas.
Estos factores, adems, pueden sufrir cambios que son transmitidos a la prole dando
lugar a la aparicin de variabilidad dentro de un mismo rasgo para individuos de una
misma especie.
Estos cambios se producen por:
Mutacin, que puede entenderse como alteracin o cambio en el mensaje
gentico de un individuo.
Recombinacin, se trata del proceso de rotura de una hembra de ADN que al
repararse se une a una molcula de ADN diferente a la que proceda.
Entrecruzamiento cromosmico, por el cual los cromosomas al aparearse
durante la meiosis intercambian fragmentos de ADN.
Instituto Hernando Arias de Saavedra
BIOLOGA
118
Aunque hoy da la palabra gen presenta mltiples significados, suele utilizarse para
referirse a los factores hereditarios.
Locus
Cromosoma.
(1) Cromtida. Una de las dos partes idnticas del cromosoma despus de la fase S. (2) Centrmero. Punto
donde las dos cromtidas contactan, y donde se unen los microtbulos. (3) Brazo corto. (4) Brazo largo.
En biologa evolutiva para identificar posiciones de inters sobre determinadas
secuencias, un locus (del latn locus, lugar; plural loci) es una posicin fija sobre un
cromosoma, como la posicin de un gen o de un biomarcador (marcador gentico). Una
variante de la secuencia de ADN en un determinado locus se llama alelo. La lista
ordenada de loci conocidos para un genoma particular se denomina mapa gentico,
mientras que se denomina cartografa gentica al proceso de determinacin del locus de
un determinado carcter biolgico.
Las clulas diploides y poliploides cuyos cromosomas tienen el mismo alelo en algn
locus se llaman homocigotos, mientras que los que tienen diferentes alelos en un locus,
heterocigotos.
Instituto Hernando Arias de Saavedra
BIOLOGA
119
Ejemplos de bandas.
Un rango de localizaciones se especifica de la misma manera. Por ejemplo, el locus del
gen OCA1 se anotara "11q1.4-q2.1", significando que est en el brazo largo del
cromosoma 11, en algn lugar del mbito de la sub-banda 4 de la banda 1 y de la sub-
banda 1 de la banda 2.
Los extremos de un cromosoma se etiquetan como "ptel" y "qtel", y as "2qtel" se refiere
al telmero del brazo largo del cromosoma 2.
Alelo
Un alelo (del griego: , alllon: uno a otro, unos a otras) es cada una de las
formas alternativas que puede tener un gen que se diferencian en su secuencia y que se
puede manifestar en modificaciones concretas de la funcin de ese gen. Al ser la mayora
de los mamferos diploides estos poseen dos alelos de cada gen, uno de ellos procedente
del padre y el otro de la madre. Cada par de alelos se ubica en igual locus o lugar del
cromosoma. Cuales alelos poseen cromosomas.
Por alelo debe entenderse el valor de dominio que se otorga a un gen cuando rivaliza
contra otro gen por la ocupacin de posicin final en los cromosomas durante la
separacin que se produce durante la meiosis celular. De ese valor de dominacin del
alelo procreador resultar la trasmisin, idntica o distinta, de la copia o serie de copias
del gen procreado. De acuerdo con esa potencia, un alelo puede ser dominante y
expresarse en consecuencia en el hijo solamente con una de las copias procreadoras,
por lo tanto si el padre o la madre lo poseen el cromosoma del hijo lo expresar siempre;
o bien puede ser un alelo recesivo, por lo tanto se necesitarn dos copias del mismo gen,
dos alelos, para que se exprese en el cromosoma procreado, esto es, deber ser provisto
al momento de la procreacin por ambos progenitores.
Instituto Hernando Arias de Saavedra
BIOLOGA
120
El concepto de alelo se entiende a partir de la palabra alelomorfo (en formas alelas) es
decir, algo que se presenta de diversas formas dentro de una poblacin de individuos.
Definicin en gentica mendeliana
Cada una de las alternativas que puede tener un gen de un carcter.
Por ejemplo, el gen que regula el color de la semilla del guisante presenta dos alelos: uno
que determina color verde y otro que determina color amarillo. Por regla general se
conocen varias formas allicas de cada gen; el alelo ms extendido de una poblacin se
denomina "alelo normal, salvaje o silvestre", mientras que los otros ms escasos, se
conocen como "alelos mutantes". As, de forma general, con dos alelos (a1 y a2)
podemos tener tres tipos de combinaciones en diploides:
Dos alelos: a
1
, a
1
(homocigoto para el alelo a
1
)
Dos alelos: a
2
, a
2
(homocigoto para el alelo a
2
)
Un alelo a
1
y otro a
2
: (heterocigoto: a
1
, a
2
)
Tipos de alelo
Los alelos son formas alternas de un gen, que difieren en secuencia o funcin.
Toda caracterstica genticamente determinada depende de la accin de cuando menos
un par de genes homlogos, que se denominan alelos.
Los alelos que varan en secuencia tienen diferencias en el ADN, como
deleciones, inserciones o sustituciones.
Los alelos que difieren en funcin pueden tener o no diferencias conocidas en las
secuencias, pero se evalan por la forma en que afectan al organismo.
En funcin de su expresin en el fenotipo se pueden dividir en:
Instituto Hernando Arias de Saavedra
BIOLOGA
121
Alelos dominantes: aquellos que aparecen en el fenotipo de los individuos
heterocigotos o hbridos para un determinado carcter, adems de en el
homocigoto.
Alelos recesivos: los que quedan enmascarados del fenotipo de un individuo
heterocigoto y slo aparecen en el homocigoto, siendo homocigtico para los
genes recesivos.
Ecuaciones
Hay dos simples ecuaciones para calcular la frecuencia de dos alelos de un gen
determinado en una poblacin de individuos (segn el principio de Hardy-Weinberg):
Ecuacin 1:p2 + 2pq + q2 = 1
Ecuacin 2: p + q = 1
Donde:
p: es la frecuencia de un alelo
q: es la frecuencia del otro alelo.
p2: es la fraccin de poblacin que es homocigtica para el alelo p.
2pq: es la frecuencia de heterocigotos
q2: es la fraccin de poblacin que es homocigtica para el alelo q.
Instituto Hernando Arias de Saavedra
BIOLOGA
122
La seleccin natural puede actuar en p y q en la ecuacin 1, cambiando su frecuencia
dentro de la poblacin de individuos; y, obviamente, afecta la frecuencia de alelos vista
en la ecuacin 2. Debera notarse que la segunda ecuacin puede derivarse de la
primera (y viceversa), dado que p2 + 2pq + q2 = 1 implica que (p + q)2 = 1 donde p y q
son nmeros positivos.
Fenotipo
Los individuos de la especie de moluscos Donax variabilis muestran
una extraordinaria diversidad fenotpica, tanto en el color como en
el patrn de sus conchas.
En biologa y ciencias de la salud, se denomina fenotipo a la expresin del genotipo en
un determinado ambiente. Los rasgos fenotpicos incluyen rasgos tanto fsicos como
conductuales. Es importante destacar que el fenotipo no puede definirse como la
"manifestacin visible" del genotipo, pues a veces las caractersticas que se estudian no
son visibles en el individuo, como es el caso de la presencia de una enzima. Un fenotipo
es cualquier caracterstica o rasgo observable de un organismo, como su morfologa,
desarrollo, propiedades bioqumicas, fisiologa y comportamiento.
El fenotipo est determinado fundamentalmente por el genotipo, o por la identidad de los
alelos, los cuales, individualmente, cargan una o ms posiciones en los cromosomas.
Algunos fenotipos estn determinados por los mltiples genes, y adems influidos por
factores del medio. De esta manera, la identidad de uno, o de unos pocos alelos
conocidos, no siempre permite una prediccin del fenotipo. En este sentido, la interaccin
entre el genotipo y el fenotipo ha sido descrita usando la simple ecuacin que se expone
a continuacin:
Ambiente + Genotipo = Fenotipo
En conclusin, el fenotipo es cualquier caracterstica detectable de un organismo
(estructural, bioqumico, fisiolgico o conductual) determinado por una interaccin entre
su genotipo y su medio.
Instituto Hernando Arias de Saavedra
BIOLOGA
123
El conjunto de la variabilidad fenotpica recibe el nombre de polifasia o polifenismo.
Determinacin del carcter color de ojos; genotipo, arriba, y fenotipo, abajo.
Dado que los fenotipos son mucho ms fciles de observar que los genotipos, la gentica
clsica usa los fenotipos para determinar las funciones de los genes. Experimentos de
reproduccin pueden probar estas interferencias. De esta forma, estudios genticos
tempranos son capaces de rastrear los patrones hereditarios sin hacer uso de la biologa
molecular.
La idea de fenotipo como el producto del genotipo ha sido generalizada por Richard
Dawkins en su libro El Fenotipo Extendido (1982).
Genotipo
El genotipo es el contenido genoma especfico de un individuo, en forma de ADN. Junto
con la variacin ambiental que influye sobre el individuo, codifica el fenotipo del individuo.
De otro modo, el genotipo puede definirse como el conjunto de genes de un organismo
y el fenotipo como el conjunto de rasgos de un organismo. Por tanto, los cientficos y
los mdicos hablan a veces por ejemplo del (gen) tipo de un cncer particular, separando
as la enfermedad del enfermo. Aunque pueden cambiar los codones para distintos
aminocidos por una mutacin aleatoria (cambiando la secuencia que codifica un gen,
eso no altera necesariamente el fenotipo. Ejemplo: si una gata blanca se cruza con un
gato negro salen gatos grises, es decir el genotipo era: gata blanca (B) gato negro (N)
son 100% heterocigotos es decir 100% grises.
Genotipo y fenotipo
Toda la informacin contenida en los cromosomas se conoce como genotipo, sin
embargo dicha informacin puede o no manifestarse en el individuo. El fenotipo se refiere
Instituto Hernando Arias de Saavedra
BIOLOGA
124
a la expresin del genotipo ms la influencia del medio. El botnico holands Wilhelm
Johannsen acu tanto el trmino gen como la distincin entre genotipo y fenotipo.
Normalmente se refiere al genotipo de un individuo con respecto a un gen de inters
particular y, en individuos poliploides, se refiere a la combinacin de los alelos que porta
el individuo (homocigoto y heterocigoto). Un cambio en un cierto gen provocar
normalmente un cambio observable en un organismo, conocido como el fenotipo. Los
trminos genotipo y fenotipo son distintos por al menos dos razones:
1. Para distinguir la fuente del conocimiento de un observador (uno puede conocer el
genotipo observando el ADN; uno puede conocer el fenotipo observando la
apariencia externa de un organismo).
2. El genotipo y el fenotipo no estn siempre correlacionados directamente. Algunos
genes solo expresan un fenotipo dado bajo ciertas condiciones ambientales.
Inversamente, algunos fenotipos pueden ser el resultado de varios genotipos. La
distincin entre genotipo y fenotipo se constata a menudo al estudiar los patrones
familiares para ciertas enfermedades o condiciones hereditarias, por ejemplo la
hemofilia. Algunas personas que no tienen hemofilia pueden tener hijos con la
enfermedad, porque ambos padres "portaban" los genes de la hemofilia en su
cuerpo, aunque stos no tenan efecto en la salud de los padres. Los padres, en
este caso, se llaman portadores. La gente sana que no es portadora y la gente
sana que es portadora del gen de la hemofilia tienen la misma apariencia externa
(es decir, no tienen la enfermedad), y por tanto se dice que tienen el mismo
fenotipo. Sin embargo, los portadores tienen el gen y el resto de la gente no
(tienen distintos genotipos).
Estudios de asociacin
Con un diseo experimental adecuado, uno puede utilizar mtodos estadsticos para
correlacionar diferencias en los genotipos de las poblaciones con diferencias en sus
fenotipos observados. Estos estudios de asociacin se pueden utilizar para determinar
los factores de riesgo asociados con una enfermedad. Pueden servir incluso para
diferenciar entre poblaciones que pueden responder favorablemente o no a un
tratamiento medicamentoso particular. Este enfoque se conoce como medicina
personalizada.
Leyes de Mendel (1865)
Leyes de Mendel
Instituto Hernando Arias de Saavedra
BIOLOGA
125
Primera ley, o Principio de la uniformidad: Cuando se cruzan dos individuos
de raza pura, los hbridos resultantes son todos iguales." individuos homocigotas,
uno dominante (AA) y otro recesivo (aa), origina slo individuos heterocigotas, es
decir, los individuos de la primera generacin filial son uniformes entre ellos (Aa).
Segunda ley, o Principio de la segregacin: Ciertos individuos son capaces de
transmitir un carcter aunque en ellos no se manifieste. El cruce de dos
individuos de la F1 (Aa) dar origen a una segunda generacin filial en la cual
reaparece el fenotipo "a", a pesar de que todos los individuos de la F1 eran de
fenotipo "A". Esto hace presumir a Mendel que el carcter "a" no haba
desaparecido, sino que slo haba sido "opacado" por el carcter "A", pero que al
reproducirse un individuo, cada carcter segrega por separado.
Conocida tambin, en ocasiones como Ley de Mendel de la segregacin
equitativa o disyuncin de los alelos. Esta ley establece que durante la formacin
Instituto Hernando Arias de Saavedra
BIOLOGA
126
de los gametos cada alelo de un par se separa del otro miembro para determinar
la constitucin gentica del gameto filial.
Mendel estudi siete caracteres que aparecen en dos formas discretas, en vez de
caracteres difciles de definir que dificultan su estudio. Lo primero que realiz
fueron cruzamientos entre plantas que diferan para slo un carcter (cruzamiento
monohbrido).
Utilizando smbolos podemos describir el cruzamiento de plantas altas x plantas enanas
de la siguiente manera:
Generacin parental: DD x dd
Gametos parentales: solamente D solamente d
Genotipo de la F1 Dd
En los cruzamientos de Mendel la segunda generacin o F2 fue obtenida por
autofecundacin de las plantas de la F1. Esto puede describirse en una tabla
denominada Tablero o cuadrado de Punnett
22
.
Tablero de Punnett
Gametos de la generacin
F1
D d
D DD (alta) Dd (alta)
d Dd (alta) dd (baja)
22 El cuadro de Punnett es un diagrama diseado por Reginald Punnett y es usado por los bilogos para
determinar la probabilidad de que un producto tenga un genotipo particular. El cuadro de Punnett permite
observar cada combinacin posible de un alelo materno con otro alelo paterno por cada gen estudiado
Instituto Hernando Arias de Saavedra
BIOLOGA
127
El tablero de Punnett nos permite determinar las proporciones genotpicas esperadas.
Tambin nos permite determinar las proporciones fenotpicas.
Proporciones Genotpicas de la F2: 1DD:2Dd:1dd
Proporciones Fenotpicas de la F2: 3 altas: 1 enana (3D_:1dd)
Cruce monohbrido clsico
En este modelo, ambos organismos poseen el genotipo Bb, por lo que pueden producir
gametos que contengan los alelos "B" y "b" (se acostumbra en los estudios de la gentica
usar maysculas para expresar los alelos dominantes y con minscula a los recesivos).
La probabilidad de que el producto tenga el genotipo BB es de 25%, con Bb es de 50% y
con bb de 25%. Todos los genotipos son alelos, por lo tanto todos son conocidos como
un punnett normal o adyacente.
Materno
B b
Paterno
B BB Bb
b Bb bb
Cabe sealar que el cuadro de Punnett solo muestra las posibilidades para genotipos, no
para fenotipos. La forma en que los alelos B y b interactan uno con el otro afectando la
apariencia del producto depende de cmo interacten los productos de los genes.
Para los genes clsicos dominantes/recesivos, como los que determinan el color del pelo
de una rata, siendo B el pelo negro y b el pelo blanco, el alelo dominante eclipsar al
recesivo.
Es muy habitual representar las posibilidades de hibridacin mediante un cuadro de
Punnett.
Cruce dihbrido clsico
Instituto Hernando Arias de Saavedra
BIOLOGA
128
Cruzamientos ms complejos pueden presentarse cuando se contemplan dos o ms
genes. El cuadro de Punnett solo funciona si los genes son independientes entre ellos.
El siguiente ejemplo ilustra un cruce dihbrido entre dos plantas heterocigticas de
guisante. R representa el alelo dominante de la forma (redondeada) mientras que r
muestra el alelo recesivo (rugoso). Y es el alelo dominante del color (amarillo) cuando y
es el alelo recesivo (verde). Si cada planta tiene el genotipo Rr Yy y los genes son
independientes, estos pueden producir cuatro tipos de gametos con todas las posibles
combinaciones: RY, Ry, rY y ry.
RY Ry rY ry
RY RRYY RRYy RrYY RrYy
Ry RRYy RRyy RrYy Rryy
rY RrYY RrYy rrYY rrYy
ry RrYy Rryy rrYy rryy
Ya que los alelos dominantes eclipsan a los recesivos hay nueve combinaciones que
tienen el fenotipo redondeado amarillo, tres que son redondeado verde, tres de
combinacin rugoso amarillo y una con el fenotipo rugoso verde. La proporcin se
muestra como 9:3:3:1 y es la ms usual para el cruce dihbrido.
EJEMPLO:
Fenotipos de las lneas puras: flor roja o blanca
Cruzamiento Parental: Rojo x Blanco
F1: Se esperara flores rojas o blancas en esta generacin dependiendo de qu alelo
fuera dominante. Pero las plantas de la F1 de este cruzamiento tenan flores rosadas. Tal
como se hara en cualquier experimento, se autofecundaron las plantas de la F1. Los
resultados obtenidos fueron:
F2: relacin fenotpica Rojas: Rosadas: Blancas
RESOLVER el ejemplo de ejercicios usando el cuadrado de Punnett para establecer las
proporciones fenotpicas y genotpicas de la descendencia de una madre rubia con ojos
azules y un padre de cabellos y ojos castaos.
Siendo:
Ojos azules: c
Ojos castaos: C
Cabello rubio: r
Cabello castao: R
Instituto Hernando Arias de Saavedra
BIOLOGA
129
Mendel obtuvo esta ley al cruzar diferentes variedades de individuos heterocigotos
(diploides con dos variantes allicas del mismo gen: Aa), y pudo observar en sus
experimentos que obtena muchos guisantes con caractersticas de piel amarilla y otros
(menos) con caractersticas de piel verde, comprob que la proporcin era de 3:4 de color
amarilla y 1:4 de color verde (3:1).
Segn la interpretacin actual, los dos alelos, que codifican para cada caracterstica, son
segregados durante la produccin de gametos mediante una divisin celular meitica.
Esto significa que cada gameto va a contener un solo alelo para cada gen. Lo cual
permite que los alelos materno y paterno se combinen en el descendiente, asegurando la
variacin.
Para cada caracterstica, un organismo hereda dos alelos, uno para cada pariente. Esto
significa que en las clulas somticas, un alelo proviene de la madre y otro del padre.
stos pueden ser homocigotos o heterocigotos.
En palabras del propio Mendel:
"Resulta ahora claro que los hbridos forman semillas que tienen el uno o el otro de
los dos caracteres diferenciales, y de stos la mitad vuelven a desarrollar la forma
hbrida, mientras que la otra mitad produce plantas que permanecen constantes y
reciben el carcter dominante o el recesivo en igual nmero. "
Gregor Mendel
Tercera ley, o Principio de la combinacin independiente: Esta ley hace
referencia al cruce polihbrido (monohbrido: cuando se considera un carcter;
polihibrido: cuando se consideran dos o ms caracteres). Mendel trabaj este
cruce en guisantes, en los cuales las caractersticas que l observaba (color de la
semilla y rugosidad de su superficie) se encontraban en cromosomas separados.
De esta manera, observ que los caracteres se transmitan
independientemente unos de otros. Esta ley, sin embargo, deja de cumplirse
cuando existe vinculacin (dos genes estn en locus muy cercanos y no se
separan en la meiosis).
En ocasiones es descrita como la 2 Ley. Mendel concluy que diferentes rasgos son
heredados independientemente unos de otros, no existe relacin entre ellos, por tanto el
patrn de herencia de un rasgo no afectar al patrn de herencia de otro. Slo se cumple
Instituto Hernando Arias de Saavedra
BIOLOGA
130
en aquellos genes que no estn ligados (en diferentes cromosomas) o que estn en
regiones muy separadas del mismo cromosoma. Es decir, siguen las proporciones
9:3:3:1.
En palabras del propio Mendel:
Por tanto, no hay duda de que a todos los caracteres que intervinieron en los
experimentos se aplica el principio de que la descendencia de los hbridos en que se
combinan varios caracteres esenciales diferentes, presenta los trminos de una serie de
combinaciones, que resulta de la reunin de las series de desarrollo de cada pareja de
caracteres diferenciales.
Gregor Mendel
Experimentos de Mendel
Mendel inici sus experimentos eligiendo dos plantas de guisantes que diferan en un
carcter, cruz una variedad de planta que produca semillas amarillas con otra que
produca semillas verdes, estas plantas forman la Generacin Parental (P).
Como resultado de este cruce se produjeron plantas que producan nada ms que
semillas amarillas, repiti los cruces con otras plantas de guisante que diferan en otros
caracteres y el resultado era el mismo, se produca un carcter de los dos en la
generacin filial. Al carcter que apareca le llamo Dominante y al que no, Recesivo. En
este caso el color amarillo es dominante frente al color verde.
Las plantas obtenidas de la Generacin Parental se denominan Primera Generacin
Filial o F1.
Mendel dej que se autofecundaran las plantas de la Primera Generacin Filial y obtuvo
la Segunda Generacin Filial o F2 compuesta por plantas que producan semillas
amarillas y plantas que producan semillas verdes en una proporcin 3:1 (3 de semillas
amarillas y 1 de semillas verdes). Repiti el experimento con otros caracteres
diferenciados y obtuvo resultados similares en una proporcin 3:1.
De esta experiencia formul la Primera y Segunda ley.
Ms adelante Mendel decidi comprobar si estas leyes funcionaban en plantas
diferenciadas en dos o ms caracteres, eligi como Generacin Parental plantas de
semillas amarillas y lisas y plantas de semillas verdes y rugosas.
Las cruz y obtuvo la Primera Generacin Filial compuesta por Plantas de semillas
amarillas y lisas, la primera ley se cumpla, en la F1 aparecan los caracteres dominantes
(amarillos y lisos) y no los recesivos (verdes y rugosos).
Instituto Hernando Arias de Saavedra
BIOLOGA
131
Obtuvo la Segunda Generacin Filial autofecundando la Primera Generacin Filial y
obtuvo semillas de todos los estilos posibles, plantas que producan semillas amarillas y
lisas, amarillas y rugosas, verdes y lisas y verdes y rugosas, las cont y prob con otras
variedades y se obtenan en una proporcin 9:3:3:1 ( 9 plantas de semillas amarillas y
lisas, 3 de semillas amarillas y rugosas, 3 de semillas verdes y lisas y una planta de
semillas verdes y rugosas).
Los siete caracteres que observ G. Mendel en sus experiencias genticas con los guisantes.
Mendel public sus experimentos con guisantes en 1865 y 1866. A continuacin se
describen las principales ventajas de la eleccin de Pisum sativum como organismo
modelo: su bajo coste, tiempo de generacin corto, elevado ndice de descendencia,
diversas variedades dentro de la misma especie (color, forma, tamao, etc.). Adems,
rene caractersticas tpicas de las plantas experimentales, como poseer caracteres
diferenciales constantes.
Pisum sativum es una planta autgama, es decir, se autofecunda. Mendel lo evit
emasculndola (eliminando las anteras). As pudo cruzar exclusivamente las variedades
deseadas. Tambin embols las flores para proteger a los hbridos de polen no
controlado durante la floracin. Llev a cabo un experimento control realizando
cruzamientos durante dos generaciones sucesivas mediante autofecundacin para
obtener lneas puras para cada carcter.
Mendel llev a cabo la misma serie de cruzamientos en todos sus experimentos. Cruz
dos variedades o lneas puras diferentes respecto de uno o ms caracteres. Como
resultado obtena la primera generacin filial (F
1
), en la cual observ la uniformidad
fenotpica de los hbridos. Posteriormente, la autofecundacin de los hbridos de F
1
dio
lugar a la segunda generacin filial (F
2
), y as sucesivamente. Tambin realiz
cruzamientos recprocos, es decir, alternaba los fenotipos de las plantas parentales:
P
1
x P
2
P
2
x P
1
Instituto Hernando Arias de Saavedra
BIOLOGA
132
(Siendo P la generacin parental y los subndices 1 y 2 los diferentes fenotipos de sta).
Adems, llev a cabo retrocruzamientos, que consisten en el cruzamiento de los hbridos
de la primera generacin filial (F
1
) por los dos parentales utilizados, en las dos
direcciones posibles:
F
1
x P
2
y P
2
x F
1
(cruzamientos recprocos)
F
1
x P
1
y P
1
x F
1
(cruzamientos recprocos)
Los experimentos demostraron que:
La herencia se transmite por elementos particulados (refutando, por tanto, la
herencia de las mezclas).
Siguen normas estadsticas sencillas, resumidas en sus dos principios.
Patrones de herencia mendeliana
Mendel describi dos tipos de "factores" (genes) de acuerdo a su expresin fenotpica en
la descendencia, los dominantes y los recesivos, pero existe otro factor a tener en cuenta
en organismos dioicos y es el hecho de que los individuos de sexo femenino tienen dos
cromosomas X (XX) mientras los masculinos tienen un cromosoma X y uno Y (XY), con lo
cual quedan conformados cuatro modos o "patrones" segn los cuales se puede trasmitir
una mutacin simple:
Gen dominante ubicado en un autosoma (herencia autosmica dominante).
Gen recesivo ubicado en un autosoma (herencia autosmica recesiva).
Gen dominante situado en el cromosoma X (herencia dominante ligada al
cromosoma X).
Gen recesivo situado en el cromosoma X (herencia recesiva ligada al cromosoma
X).
Fenmenos que alteran las segregaciones mendelianas
Herencia ligada al sexo
Es la herencia con el par sexual. El cromosoma X porta numerosos genes en tanto el
cromosoma Y tan solo unos pocos y la mayora en relacin con la masculinidad. El
cromosoma X es comn para ambos sexos, pero solo el hombre posee cromosoma Y.
Instituto Hernando Arias de Saavedra
BIOLOGA
133
Herencias influidas por el sexo y limitadas al sexo
En las herencias limitadas al sexo pueden estar comprometidas mutaciones de genes
con cromosomas autosmicos cuya expresin solamente tiene lugar en rganos del
aparato reproductor masculino o femenino. Un ejemplo es el defecto congnito septum
vaginal transverso, de herencia autosmica recesiva, o la deficiencia de 5 reductasa
que convierte a la testosterona en dihidrotestosterona que acta en la diferenciacin de
los genitales externos masculinos, por lo que su ausencia simula genitales femeninos
cuando el nio nace.
Una mutacin puede estar influida por el sexo, esto puede deberse al efecto del
metabolismo endocrino que diferencia a machos y hembras. Por ejemplo, en humanos la
calvicie se debe al efecto de un gen que se expresa como autosmico dominante, sin
embargo en una familia con la segregacin de este gen solo los hombres padecen de
calvicie y las mujeres tendrn su cabello ms escaso despus de la menopausia. Otro
ejemplo puede ser la deficiencia de la enzima 21 hidroxilasa que interviene en el
metabolismo de los glucocorticoides. Cuando esta enzima est ausente, la sntesis de
glucocorticoides se desplaza hacia la formacin de testosterona y esta hormona est
comprometida en la embriognesis de los genitales externos del varn, por lo que su
presencia anormal en el desarrollo de un feto femenino produce la masculinizacin de los
genitales femeninos, mientras que en el caso de un feto varn, solo incrementa el
desarrollo de los masculinos. Una anormalidad de este tipo, permitir sospechar un
diagnostico clnico ms rpidamente en una nia, basado en el examen de los genitales
del recin nacido, que en un nio.
Instituto Hernando Arias de Saavedra
BIOLOGA
134
Estructura gnica del cromosoma Y
Por tener un solo cromosoma X, a los individuos de sexo masculino no se les pueden
aplicar los trminos "homocigoto" o "heterocigoto" para genes ubicados en este
cromosoma y ausentes en el cromosoma Y. Ya sean genes que expresen el carcter
dominante o recesivo, si estn situados en el cromosoma X, los varones siempre lo
expresarn y al individuo que lo porta se le denomina hemicigoto.
De lo anterior se deduce que, puesto que las hembras tienen un solo tipo de cromosoma
sexual, el X, sus gametos siempre tendrn la dotacin cromosmica 23,X, mientras los
masculinos pueden portar una X, dando lugar a un individuo femenino (XX), o una Y, con
lo que se originara un individuo masculino (XY). Debido a esto se dice que las mujeres
son homogamticas (todos sus gametos tienen igual constitucin) y que los hombres son
heterogamticos (tienen gametos 23,X y 23,Y).
Sistema de compensacin de dosis gnica del cromosoma X
En insectos, tal como se ha visto en Drosophila, se descubri la existencia de un gen que
ejerce de compensador de dosis, cuando se encuentra en dosis nica (como ocurre en
machos) produce la activacin de la expresin de los genes del cromosoma X. En
mamferos no se ha encontrado un gen con funcin equivalente.
Lionizacin
La lionizacin o inactivacion del cromosoma X se produce porque, a diferencia del
cromosoma Y, el X tiene gran cantidad de genes activos que codifican para importantes
productos, tales como el factor VIII de la coagulacin. Podra pensarse, por tanto, que si
las hembras tienen dos X deben tener el doble de los productos o enzimas cuyos genes
estn en ese cromosoma con relacin a los individuos del sexo masculino, sin embargo,
esto no ocurre as.
Se ha observado en mamferos que en las clulas somticas del sexo femenino (46,XX),
solo uno de los dos cromosomas X es activo. El otro permanece inactivo y aparece en
clulas en interfase como un cuerpo denso fuertemente coloreado, que se inactiva y se
adosa a la membrana nuclear en la periferia del ncleo, y que recibe el nombre de cuerpo
de Barr. La inactivacin del cromosoma X tiene lugar en el estado de mrula, alrededor
del tercer da despus de la fertilizacin y se completa, en la masa de clulas internas
que darn origen al embrin, al final de la primera semana de desarrollo embrionario. La
seleccin del cromosoma X que se inactivar, es un fenmeno generalmente aleatorio
teniendo en cuenta que al ocurrir la fecundacin cada cromosoma X tiene origen materno
y paterno, en unas clulas se inactivar el X materno (Xm) y en otras el X paterno (Xp).
Una vez que se inactiva uno de los dos cromosomas X las clulas descendientes
mantendrn el mismo cromosoma X inactivo originndose un clon celular (Xm) o (Xp)
activos. Es decir, al inicio de la inactivacin, sta es al azar, primero se inactiva al azar
cualquiera de las dos X, ya sea la heredada de la madre o del padre; pero una vez
Instituto Hernando Arias de Saavedra
BIOLOGA
135
ocurrida se mantiene el mismo cromosoma X que se inactiv en la primera clula del clon
y las clulas que deriven de sta durante el proceso de crecimiento y desarrollo
mantendrn en adelante inactivado el mismo cromosoma X.
La inactivacin (desactivacin) del cromosoma X est determinada por el gen XIST. Este
gen est involucrado en la transcripcin especfica de inactivacin que funciona por un
mecanismo de metilacin preferencial, esto significa que si no hay ninguna alteracin de
estructura en los dos cromosomas X del genoma femenino, la inactivacin debe ocurrir
de forma aleatoria, pero si existiera alguna alteracin con gran compromiso en la funcin
de uno de los dos cromosomas X habra una activacin no completamente aleatoria. El
locus del gen XIST se encuentra localizado en Xq13.3.
La inactivacin del X determina consecuencias genticas y clnicas:
Compensacin de dosis: iguala la dosis de productos de genes con el
hemicigtico para genes localizados en el cromososa X, determinando
concentraciones proteicas similares en ambos sexos, para genes ligados al X.
Variaciones en la expresin de mutaciones en hembras heterocigticas: por
ejemplo, presencia de sntomas ms o menos severos en hembras portadoras
para hemofilias A o B, distrofia muscular Duchenne, distrofias retinianas recesivas
ligadas al X.
Los rganos femeninos se comportan como mosaicos. Este fenmeno se puede
manifestar en zonas en las que se manifieste un alelo (procedente del X de la
madre) y otras zonas en las que se manifiesta el otro alelo. Se observa en
fenmenos como el color del pelaje de algunas hembras de felinos, de forma que
los felinos de tres colores son hembras, y los de dos colores son machos; en el
albinismo ocular recesivo ligado al X; o en el test inmunohistoqumico para la
deteccin de la distrofina en hembras heterocigticas para la distrofia muscular
Duchenne.
Penetrancia de un gen o de una mutacin especifica
Penetrancia es el trmino que se emplea para referirse a la expresin en trminos de
todo o nada dentro de una poblacin de individuos. Si la mutacin se expresa en menos
del 100% de los individuos portadores o heterocigticos se dice que la mutacin tiene una
penetrancia reducida y que ese individuo aparentemente sano para el carcter o
enfermedad que se estudia en la familia puede trasmitir la mutacin a su descendencia y
stos expresar el defecto. La penetrancia reducida parece ser el efecto de la relacin de
la mutacin en cuestin y otros genes del genoma, con los cuales se encuentra
interactuando.
Expresividad de un gen o mutacin especifica
Expresividad se usa para referirse al grado de severidad que se manifiesta en el fenotipo.
En trminos clnicos, es sinnimo de gravedad. La expresin de un gen tambin depende
Instituto Hernando Arias de Saavedra
BIOLOGA
136
de la relacin de ste con el resto del genoma, pero tambin de la relacin genoma-
ambiente. Para referirse a estas gradaciones fenotpicas se utiliza el trmino expresividad
variable del gen o de la mutacin.
Efecto pleiotrpico de un gen o mutacin especifica
Con el trmino pleiotropa o efecto pleiotrpico de un gen se hace referencia a todas
las manifestaciones fenotpicas en diferentes rganos o sistemas que son explicables por
una simple mutacin. Un ejemplo clsico para explicar este trmino lo constituye el
sndrome Marfan, cuya mutacin afecta al gen FBN1 que codifica a la protena fibrilina,
esta protena se encuentra en el tejido conectivo y explica las manifestaciones
esquelticas, oculares y cardiovasculares que caracterizan al sndrome.
Heterogeneidad gentica
Este trmino se aplica tanto a mutaciones en genes localizados en diferentes
cromosomas que producen expresin similar en el fenotipo (heterogeneidad no allica)
como a mutaciones que afectan a diferentes sitios del mismo gen (heterogeneidad
allica). Esta categora complica extraordinariamente el estudio etiolgico de variantes
del desarrollo de origen gentico y constituye una amplia y fundamental fuente de
diversidad gentica del desarrollo.
Nuevas mutaciones con expresin dominante
Cuando tiene lugar una mutacin de novo que se expresa como dominante, o sea, en un
genotipo heterocigtico, ocurre que padres que no presentan el efecto de la mutacin
pueden tener un descendiente afectado. La ausencia de antecedentes familiares, una vez
que se excluyen fenmenos como la penetrancia reducida del gen y variaciones mnimas
de la expresividad dificulta llegar al planteamiento de una mutacin de novo cuando en la
literatura el defecto o enfermedad no ha sido reportada con anterioridad, con un tipo
especfico de herencia.
Efecto de letalidad en un genotipo especifico
Algunas mutaciones se expresan de forma tan severa que producen letalidad en un
genotipo especfico. Un ejemplo pudiera ser el efecto de una doble dosis de una mutacin
que se expresa como dominante o el efecto en un genotipo hemicigtico, como ocurre en
la incontinencia pigmenti, enfermedad humana dominante ligada al cromosoma X.
Instituto Hernando Arias de Saavedra
BIOLOGA
137
Herencia en mamferos
El rbol genealgico
Pedigree autosmico dominante.
Como en cualquier otra especialidad mdica, en gentica adquiere enorme importancia el
interrogatorio del individuo enfermo y sus familiares, pero, adicionalmente, es vital
establecer los lazos de parentesco entre los individuos afectados y los supuestamente
sanos, por eso se utiliza el llamado rbol genealgico o pedigree en el que mediante
smbolos internacionalmente reconocidos se describe la composicin de una familia, los
individuos sanos y enfermos, as como el nmero de abortos, fallecidos, etc.
Herencias dominantes
Cuando el gen productor de una determinada caracterstica (o enfermedad) se expresa
an estando en una sola dosis se denomina dominante y los linajes donde se segrega
muestran un rbol genealgico en que, como regla, hay varios individuos que lo expresan
y los afectados tienen un progenitor igualmente afectado. No obstante, hay diferencias de
acuerdo a si el gen est ubicado en un autosoma o en el cromosoma X.
En la herencia autosmica dominante se cumplen los siguientes hechos:
Varios individuos afectados.
Los afectados son hijos de afectados.
Se afectan por igual hombres y mujeres.
Como regla, la mitad de la descendencia de un afectado hereda la afeccin.
Los individuos sanos tienen hijos sanos.
Instituto Hernando Arias de Saavedra
BIOLOGA
138
Hay hombres afectados hijos de hombres afectados (lo cual excluye la posibilidad
de que el gen causante de la afeccin est ubicado en el cromosoma X, que en
los varones procede de la madre).
El patrn ofrece un aspecto vertical.
En este caso los individuos afectados son usualmente heterocigticos y tienen un riesgo
del 50% en cada intento reproductivo de que su hijo herede la afeccin
independientemente de su sexo.
En la herencia dominante ligada al cromosoma X, aunque el gen sea dominante, si est
ubicado en el cromosoma X, el rbol genealgico suele mostrar algunas diferencias con
respecto al de la herencia autosmica dominante:
Aunque los afectados usualmente son hijos de afectados y la mitad de la
descendencia presenta la afeccin, no podemos identificar varones que hayan
heredado la afeccin de su padre, o sea, no hay trasmisin varn-varn, puesto
que los padres dan a sus hijos el cromosoma Y.
Igualmente llama la atencin que hay un predominio de mujeres afectadas pues
mientras estas pueden heredar el gen de su madre o de su padre, los varones
slo lo adquieren de su madre.
Una mujer afectada tendr el 50% de su descendencia afectada, mientras que el
hombre tendr 100% de hijas afectadas y ningn hijo afectado.
Herencias recesivas
Cuando el gen causante de la afeccin es recesivo, por regla general el nmero de
afectados es mucho menor y suele limitarse a la descendencia de una pareja, pero es
ms evidente la diferencia en la trasmisin segn la mutacin est situada en un
autosoma o en el cromosoma X.
En la herencia autosmica recesiva llama la atencin la aparicin de un individuo
afectado fruto de dos familias sin antecedentes. Esto ocurre pues ambos padres de este
individuo son heterocigticos para la mutacin, la cual, por ser recesiva, no se expresa ya
que existe un alelo dominante normal, pero, como estudiamos en las leyes de Mendel,
existe un 25% en cada embarazo, de que ambos padres trasmitan el alelo mutado,
independientemente del sexo del nuevo individuo. Por aparecer usualmente en la
descendencia de un matrimonio, se dice que su patrn es horizontal. Otro aspecto a
sealar es que cuando existe consanguinidad, aumenta la probabilidad de aparicin de
este tipo de afecciones, debido a que ambos padres comparten una parte de su genoma
proporcional al grado de parentesco entre ellos.
En la herencia recesiva ligada al cromosoma X es evidente que los individuos afectados
son todos del sexo masculino; esto se justifica porque al tener la mujer dos X y ser el gen
recesivo, el alelo dominante normal impide su expresin, mientras el varn hemicigtico
si tiene la mutacin la expresar. Tambin se observa que entre dos varones afectados
Instituto Hernando Arias de Saavedra
BIOLOGA
139
existe una mujer, que en este caso es portadora de la mutacin. La probabilidad de
descendencia afectada depender del sexo del progenitor que porta la mutacin:
Un hombre enfermo tendr 100% de hijas portadoras y 100% de hijos sanos.
Una mujer portadora tendr 50% de sus hijas portadoras y 50% de hijos varones
enfermos.
Conclusiones y comentarios
Es un error muy extendido suponer que la uniformidad de los hbridos que Mendel
observ en sus experimentos es una ley de transmisin, pues la dominancia nada tiene
que ver con la transmisin, sino con la expresin del genotipo. Por lo que esta
observacin mendeliana no suele considerarse una ley. Las leyes mendelianas de
transmisin son por lo tanto dos: la Ley de segregacin de caracteres independientes (1
ley) y la Ley de la herencia independiente de caracteres (2 ley).
Recorriendo la web de Wikipedia se puede observar este hecho, por ejemplo, en las
versiones inglesa, francesa y portuguesa consideran que las Leyes de Mendel son dos.
En cambio, en otras versiones como la catalana, la alemana, la italiana y la vasca siguen
considerando la Ley de la Uniformidad como la primera Ley de Mendel, sin ser
estrictamente una ley de transmisin de caracteres.
Incluso a nivel docente y bibliogrfico sigue permaneciendo vigente esta visin que
debera ser aclarada. Factor mendeliano: El concepto de factor mendeliano fue
introducido en 1860 por Mendel, actualmente denominado gen, ste se puede definir
como una unidad fsica y funcional que ocupa una posicin especfica en el genoma.
Lectura
Gregor Mendel
Gregor Johann Mendel-Angustuos. Naci el 20 de julio de 1822 en Hynice, Imperio
austraco y muere a los 61 aos, el 6 de enero de 1884 en Brno, Austria-Hungra. Fue un
Instituto Hernando Arias de Saavedra
BIOLOGA
140
monje agustino catlico y naturalista, nacido en Heinzendorf, Austria (actual Hynice,
distrito Nov Jin, Repblica Checa), que describi las llamadas Leyes de Mendel que
rigen la herencia gentica, por medio de los trabajos que llev a cabo con diferentes
variedades de la planta del guisante (Pisum sativum). Los primeros trabajos en gentica
fueron realizados por Mendel. Inicialmente realiz cruces de semillas, las cuales se
particularizaron por salir de diferentes estilos y algunas de su misma forma. En sus
resultados encontr caracteres como los dominantes que se caracterizan por determinar
el efecto de un gen y los recesivos por no tener efecto gentico sobre una persona
heterocigtica.
Su trabajo no fue valorado cuando lo public en el ao 1866. Hugo de Vries, botnico
holands, junto a Carl Correns y Erich von Tschermak, redescubrieron las leyes de
Mendel por separado en el ao 1900.
Biografa
Mendel naci en un pueblo llamado Heinzendorf, perteneciente al Imperio austrohngaro
(hoy Hynice, en el norte de Moravia, Repblica Checa) el 20 de julio de 1822, siendo
bautizado con el nombre de Johann Mendel. Tom el nombre de padre Gregorio al
ingresar como fraile agustino en 1843, en el convento de agustinos de Brno (conocido en
la poca como Brnn). En 1847 se orden sacerdote.
Mendel fue titular de la prelatura de la Imperial y Real Orden Austriaca del emperador
Francisco Jos I, director emrito del Banco Hipotecario de Moravia, fundador de la
Asociacin Meteorolgica Austriaca, miembro de la Real e Imperial Sociedad Morava y
Silesia para la Mejora de la Agricultura, Ciencias Naturales y Conocimientos del Pas, y
jardinero (de hecho aprendi de su padre como hacer injertos y cultivar rboles frutales).
Mendel present sus trabajos en las reuniones de la Sociedad de Historia Natural de
Brnn, (Brno), el 8 de febrero y el 8 de marzo de 1865, publicndolos posteriormente
como Experimentos sobre hibridacin de plantas (Versuche ber Plflanzenhybriden) en
1866 en las actas de la Sociedad. Sus resultados fueron ignorados por completo, y
tuvieron que transcurrir ms de treinta aos para que fueran reconocidos y entendidos.
Al tipificar las caractersticas fenotpicas (apariencia externa) de los guisantes las llam
caracteres. Us el nombre de elemento, para referirse a las entidades hereditarias
separadas. Su mrito radica en darse cuenta de que en sus experimentos (variedades de
guisantes) siempre ocurran en variantes con proporciones numricas simples.
Los elementos y caracteres han recibido posteriormente infinidad de nombres, pero
hoy se conocen de forma universal por la que sugiri en 1909 el bilogo dans Wilhem
Ludwig Johannsen, como genes. Siendo ms exactos, las versiones diferentes de genes
responsables de un fenotipo particular, se llaman alelos. Los guisantes verdes y amarillos
corresponden a distintos alelos del gen responsable del color.
Mendel falleci el 6 de enero de 1884 en Brnn, por nefritis crnica.
Instituto Hernando Arias de Saavedra
BIOLOGA
141
ANEXO
Mendel y la apicultura
Escultura de Mendel en el jardn del convento de Brno en el que
investig.
Un aspecto no muy conocido de la vida de Mendel fue que se dedic durante los ltimos
10 aos de su vida a las abejas. Mendel reconoce que las abejas result un modelo de
investigacin frustrante. Es probable que el experimento realizado con abejas fuera
guiado para confirmar la teora de la herencia.
Uno puede presumir que en 1854 Mendel discute en Silesia con los apicultores la
hiptesis de Jan Dzierzon que enuncia que las reinas infrtiles o los huevos que no son
fecundados por esperma de los machos producen znganos, producindose
reproduccin sexual en las hembras y reproduccin asexual en los machos o znganos.
A este proceso Jan Dzierzon lo denomin partenognesis.
La teora de Dzierzon fue confirmada por hibridacin, si bien el cruce de abejas es difcil,
pues durante el vuelo nupcial de la reina no debe haber znganos extraos. Por ello,
Mendel construy una jaula de tejido de cuatro metros de largo y cuatro de alto, situando
la colmena en el exterior de ella, para lograr el objetivo deseado que era realizar los
cruces necesarios para lograr los hbridos de diferentes razas de abejas. Pero durante la
vida de Mendel la teora de Dzierzon no fue probada. Seguramente, lo que Mendel
pretenda era probar la segregacin de caracteres genticos.
El director de la Sociedad de Apicultura de Brnn (Brno), Ziwansky, provey diferentes
razas de abejas de la especie Apis mellifera: italianas (Apis mellifera ligustica), carniolas
(Apis mellifera carnica), egipcias y chipriotas, que los apicultores locales reproducan. Las
chipriotas fueron obtenidas directamente de Chipre por el conde Kolowrat. Unas de las
abejas con diferencias de colores fueron obtenidas de Pernambuco (Brasil), incluyendo
algunos especmenes de Sudamrica. Estos fueron enviados por el profesor Macowsky a
Instituto Hernando Arias de Saavedra
BIOLOGA
142
Mendel y eran abejas de la especie Trigona lineata, melipnidos o abejas sin aguijn,
criadas durante dos aos sucesivos.
Mendel fue un activo miembro de la Sociedad de Apicultura de Brnn (Brno) y en 1871
fue nombrado presidente de la misma. Entre el 12 y el 14 de septiembre de 1871, Mendel
y Ziwansky fueron delegados por la Asociacin de Apicultura de Brnn (Brno) al
Congreso de Apicultura en lengua germana a desarrollarse en Kiel. En 1873 Mendel
declin la presidencia y en 1874 fue reelecto pero por circunstancias personales privadas
indic que le resultaba imposible ocupar el cargo. En 1877 se afirma en Honigbienen (la
revista de la Asociacin) que el prelado de las abejas posea 36 colmenas. Pero en
realidad el inters biolgico de Mendel resida en la relacin que tienen las abejas con las
flores.
En Europa fue William Bateson, quien impuls en 1900 el conocimiento de las leyes de
Mendel. Al dar una conferencia en la Sociedad de Horticultura, tuvo conocimiento del
trabajo de Mendel, a travs del relato de Hugo de Vries; as encontr el refrendo de lo
que haba estado experimentando. l fue, pues, quien dio las primeras noticias en
Inglaterra de las investigaciones de Mendel. En 1902, public Los principios mendelianos
de la herencia: una defensa acompaada de la traduccin de los trabajos originales de
Mendel sobre hibridacin. Adems, fue el primero en acuar trminos como "gentica",
"gen" y "alelo" para describir muchos de los resultados de esta nueva ciencia biolgica.
En 1902, Theodore Boveri y Walter Sutton, trabajando de manera independiente, llegaron
a una misma conclusin y propusieron una base biolgica para los principios
mendelianos, denominada Teora cromosmica de la herencia. Esta teora sostiene que
los genes se encuentran en los cromosomas y al lugar cromosmico ocupado por un gen
se le denomin locus (se habla de loci si se hace referencia al lugar del cromosoma
ocupado por varios genes). Ambos se percataron de que la segregacin de los factores
mendelianos (alelos) se corresponda con la segregacin de los cromosomas durante la
divisin meitica (por tanto, exista un paralelismo entre cromosomas y genes).
Algunos trabajos posteriores de bilogos y estadsticos tales como R.A. Fisher (1911)
mostraron que los experimentos realizados por Mendel tenan globalidad en todas las
especies, mostrando ejemplos concretos de la naturaleza. Los principios de la
segregacin equitativa (2 ley de Mendel) y la transmisin independiente de la herencia
(3 ley de Mendel) derivan de la observacin de la progenie de cruzamientos genticos,
no obstante, Mendel no conoca los procesos biolgicos que producan esos fenmenos.
As, puede considerarse que las leyes de Mendel reflejan el comportamiento
cromosmico durante la meiosis: la primera ley responde a la migracin aleatoria de los
cromosomas homlogos a polos opuestos durante la anafase I de la meiosis (tanto los
alelos como los cromosomas homlogos segregan de manera equitativa o 1:1 en los
gametos) y la segunda ley, al alineamiento aleatorio de cada par de cromosomas
homlogos durante la metafase I de la meiosis (por lo que genes distintos y pares
diferentes de cromosomas homlogos segregan independientemente).
Instituto Hernando Arias de Saavedra
BIOLOGA
143
Honores
- En 1994, la Universidad Mendel pasa a llamarse con su nombre en su honor.
- Un colegio mayor de la Universidad de Madrid lleva su nombre
Glosario
Gen: Es una regin de DNA que codifica para RNA.
Genotipo: factores hereditarios internos de un organismo, sus genes y por extensin su
genoma.
Fenotipo: las cualidades fsicas observables en un organismo, incluyendo su
morfologa, fisiologa y conducta a todos los niveles de descripcin.
Alelo: Es cada una de las variantes de un locus. Cada alelo aporta diferentes
variaciones al carcter que afecta. En organismos diploides (2n) los alelos de un mismo
locus se ubican fsicamente en los pares de cromosomas homlogos.
Locus: Ubicacin del gen en un cromosoma. Para un locus puede haber varios alelos
posibles. (Plural: LOCI)
Cariotipo: Composicin fotogrfica de los pares de cromosomas de una clula,
ordenados segn un patrn estndar. En un cariotipo encontramos el conjunto de
caractersticas que permiten reconocer la dotacin cromosmica de una clula.
Lnea pura: Es la descendencia de uno o ms individuos de constitucin gentica
idntica, obtenindose por autofecundacin o cruces endogmicos. Son individuos
homocigotos para todos sus caracteres.
Autofecundacin: Proceso de reproduccin sexual donde los gametos masculinos de
un individuo se fecundan con los vulos del mismo individuo. Es indispensable que sean
especies monoicas (caracterstico de las plantas y algunos animales inferiores).
Dominancia, Alelo dominante: Predominio de la accin en un alelo sobre la de su
alternativo (llamado alelo recesivo), enmascarando u ocultando sus efectos. El carcter
hereditario dominante es el que se manifiesta en el fenotipo (conjunto de las
propiedades manifiestas en un individuo). Segn la terminologa mendeliana se expresa
como A>a (el alelo A domina sobre el alelo a, el carcter que determina, es por tanto el
que observaremos en el fenotipo).
Recesividad, Alelo recesivo: Caracterstica del alelo recesivo de un gen que no se
manifiesta cuando est presente el alelo dominante. Para que este alelo se observe en
el fenotipo, el organismo debe poseer dos copias del mismo alelo, es decir, debe ser
homocigoto para ese gen (segn la terminologa mendeliana, se expresara como aa).
Instituto Hernando Arias de Saavedra
BIOLOGA
144
Meiosis: La meiosis es el proceso de divisin celular que permite a una clula diploide
generar clulas haploides en eucariotas. En este proceso se produce una replicacin
del DNA (en la fase S) y dos segregaciones cromosmicas, de manera que de una
clula inicial diploide se obtienen cuatro clulas haploides.
Homocigoto: Individuo puro para uno o ms caracteres, es decir, que en ambos loci
posee el mismo alelo (representado como aa en el caso de ser recesivo o AA si es
dominante).
Heterocigoto: Individuo que para un gen, tiene un alelo distinto en cada cromosoma
homlogo. Su representacin mendeliana es Aa.
Hbrido: Es el resultado del cruzamiento o apareamiento de dos individuos puros
homocigotos (uno de ellos recesivo y el otro dominante) para uno o varios caracteres.
Gameto: Clula sexual que procede de una estirpe celular llamada lnea germinal, en
los seres superiores tienen un nmero de cromosomas haploide (n) debido a un tipo de
divisin celular llamado meiosis que permite reducir el nmero de cromosomas a la
mitad. El gameto femenino se denomina vulo; el gameto masculino recibe el nombre
de espermatozoide.
Cigoto o huevo: Clula resultante de la unin de dos gametos haploides (es por tanto,
diploide, 2n). Generalmente, experimenta una serie de divisiones celulares hasta que se
constituye en un organismo completo. Su citoplasma y sus orgnulos son siempre de
origen materno al proceder del vulo.
Haploide: Que posee un solo juego de cromosomas (n), caracterstico de los gametos
eucariotas y los gametofitos de las plantas.
Diploide: Que tiene doble juego de cromosomas (2n). Caractersticas de las clulas
somticas.
Autosoma: Todo cromosoma que no sea sexual.
Bibliografa
GRIFFITHS, A.J.F., S. R. WESSLER, R.C. LEWONTIN & S. B. CARROLL (2008).
Gentica. MGraw-Hill Interamericana. Novena edicin.
KLUG, W.S. & CUMMINGS, M.R. (1.998). Conceptos de Gentica. 5 Edicin. Prentice
Hall. Espaa.
BENITO-JIMENEZ, C. (1.997). 360 Problemas de Gentica. Resueltos paso a paso. 1
Edicin. Editorial Sntesis. Espaa.
MENSUA, J.L. (2002). Gentica: Problemas y ejercicios resueltos. Prentice
Anthony J.F. Griffiths, Montserrat Elas Arnanz (2002). Gentica, 3 edicin, McGraw-Hill
Interamericana.
Klug, W. S., et al, Conceptos de Gentica, 8 ed. Pearson Educacin, 2006
Vous aimerez peut-être aussi
- Envoltura NuclearDocument4 pagesEnvoltura NuclearAiramTenayPas encore d'évaluation
- TEMA6 Biologia de La CelulaDocument10 pagesTEMA6 Biologia de La CelulaClaudia Aldrey GolmarPas encore d'évaluation
- Resumen Nucleo EYFAMDocument8 pagesResumen Nucleo EYFAMBbm NosbornPas encore d'évaluation
- Unidad III Núcleo CelularDocument4 pagesUnidad III Núcleo CelularRoanna BacaPas encore d'évaluation
- Núcleo CelularDocument10 pagesNúcleo Celularfunweb100% (1)
- Genética SeminarioDocument7 pagesGenética SeminarioMayerling V. RiosPas encore d'évaluation
- Ensayo-El Nucleo CelularDocument18 pagesEnsayo-El Nucleo CelularDarwin TapuyoPas encore d'évaluation
- Organelas Celulares: Núcleo y Ácidos NucleicosDocument17 pagesOrganelas Celulares: Núcleo y Ácidos NucleicosMarcela BarriosPas encore d'évaluation
- Compartimientos celulares: El núcleo celularDocument16 pagesCompartimientos celulares: El núcleo celularAlessandra CancemiPas encore d'évaluation
- Semana 7 - TeoriaDocument23 pagesSemana 7 - TeoriajoselinPas encore d'évaluation
- Actividad 5 de Morfo Adriana BritoDocument14 pagesActividad 5 de Morfo Adriana BritoAdriana BritoPas encore d'évaluation
- Atlas Celula 04 NucleoDocument12 pagesAtlas Celula 04 NucleoKelvin SosaPas encore d'évaluation
- Eucariotas: La vida complejaDocument5 pagesEucariotas: La vida complejagenialmPas encore d'évaluation
- N7 Ciclo CelularDocument8 pagesN7 Ciclo CelularKathia Meneses VioletaPas encore d'évaluation
- Ultra Estructura Del Núcleo de La CélulaDocument13 pagesUltra Estructura Del Núcleo de La CélulaEduardo Yamunaqué CastroPas encore d'évaluation
- TEMA 29 NucleoDocument16 pagesTEMA 29 NucleoJose FrcoPas encore d'évaluation
- Semana 7 Teoria - Nucleo CelularDocument17 pagesSemana 7 Teoria - Nucleo CelularEliza huamaniPas encore d'évaluation
- Taller Núcleo CelularDocument9 pagesTaller Núcleo CelularNOAM HERNANDO BALDOVINO TORRESPas encore d'évaluation
- Núcleo Celular PDFDocument38 pagesNúcleo Celular PDFBety MartínPas encore d'évaluation
- Características del núcleo interfásicoDocument7 pagesCaracterísticas del núcleo interfásicoalbert einsteinPas encore d'évaluation
- El Núcleo de La CélulaDocument16 pagesEl Núcleo de La CélulaCy MnPas encore d'évaluation
- Tema 19 Nucleo InterfasicoDocument12 pagesTema 19 Nucleo Interfasicochristina8reales8ca8Pas encore d'évaluation
- Núcleo Celular 2023Document45 pagesNúcleo Celular 2023José Javier Argüello castillo APas encore d'évaluation
- Tema 9. El NúcleoDocument12 pagesTema 9. El NúcleoMarco MatellánPas encore d'évaluation
- EstructuraNúcleoDocument37 pagesEstructuraNúcleoBety MartínPas encore d'évaluation
- Envoltura NuclearDocument12 pagesEnvoltura NuclearBikner Leon ParedesPas encore d'évaluation
- Nucleo InterfasticoDocument18 pagesNucleo InterfasticoAnahi Alban100% (1)
- Fisiologia y Morfologia Del Núcleo CelularDocument13 pagesFisiologia y Morfologia Del Núcleo CelularAbelardo Ortiz50% (4)
- El Núcleo Interfásico y El Núcleo en División.Document18 pagesEl Núcleo Interfásico y El Núcleo en División.ElenaPas encore d'évaluation
- Dr. Eduardo Kremenchutzky - Introducción al núcleo celularDocument26 pagesDr. Eduardo Kremenchutzky - Introducción al núcleo celularAlexka Silva CamachoPas encore d'évaluation
- Tecnico en Emergencias Sanitarias Trimestre 1 Anatomofisiología Con Ejercicios ResueltosDocument26 pagesTecnico en Emergencias Sanitarias Trimestre 1 Anatomofisiología Con Ejercicios ResueltosJosé María Guerrero García100% (1)
- Capítulo 4 - Núcleo CelularDocument36 pagesCapítulo 4 - Núcleo CelularKaren Velázquez BravoPas encore d'évaluation
- TEMA 29. Núcleo Interfásico y Núcleo en DivisiónDocument18 pagesTEMA 29. Núcleo Interfásico y Núcleo en DivisiónTete TetPas encore d'évaluation
- El núcleo interfásico: estructura y funcionesDocument7 pagesEl núcleo interfásico: estructura y funcionesGkfi XjdjPas encore d'évaluation
- Bloque IIIDocument251 pagesBloque IIIGuillermo MazariegosPas encore d'évaluation
- Semana 7 El Núcleo, Teoria 23-I PDFDocument35 pagesSemana 7 El Núcleo, Teoria 23-I PDFVALERYROSITA ATENCIOASCAMAPas encore d'évaluation
- HISTOLOGIA3Document11 pagesHISTOLOGIA3aarleth.aguilarPas encore d'évaluation
- Ingreso-22na771 Cleo Interfa771 Sico 2Document14 pagesIngreso-22na771 Cleo Interfa771 Sico 2Nicolas Mota FrançaPas encore d'évaluation
- Núcleo y NucléoloDocument36 pagesNúcleo y NucléoloAmilca R Trejo CPas encore d'évaluation
- TEMA 4 NucleoDocument8 pagesTEMA 4 NucleoALEX BERMUDOPas encore d'évaluation
- B2 T5 - CÉLULA EUCARIOTA - Núcleo ApuntesDocument7 pagesB2 T5 - CÉLULA EUCARIOTA - Núcleo ApuntesNacho Orio FernándezPas encore d'évaluation
- Tema 06 - Núcleo Celular.Document19 pagesTema 06 - Núcleo Celular.Saul TamayoPas encore d'évaluation
- Nucleo CelularDocument23 pagesNucleo CelularMisael VargasPas encore d'évaluation
- Tarea Semana 4Document7 pagesTarea Semana 4Yuneydis GuzmanPas encore d'évaluation
- Tema 2.nivel CelularDocument66 pagesTema 2.nivel CelularcesarPas encore d'évaluation
- Nucleo Cromosomas y GenesDocument37 pagesNucleo Cromosomas y GenesJorge Luis Montaño GutiérrezPas encore d'évaluation
- Estructura y funciones del núcleo celularDocument29 pagesEstructura y funciones del núcleo celularRubí RecinosPas encore d'évaluation
- TP - Ciclo Celular y Núcleo CelularDocument6 pagesTP - Ciclo Celular y Núcleo CelularvirginiaPas encore d'évaluation
- Libro. Neurona (Biopsicología)Document12 pagesLibro. Neurona (Biopsicología)Jess Lara McDermidPas encore d'évaluation
- TEMA 10 - El Núcleo InterfásicoDocument2 pagesTEMA 10 - El Núcleo InterfásicoOlimpia Aroca GutierrezPas encore d'évaluation
- TEMA 29.-Núcleo Interfásico, Núcleo en DivisiónDocument17 pagesTEMA 29.-Núcleo Interfásico, Núcleo en DivisiónRu DSPas encore d'évaluation
- 3 NúcleoDocument10 pages3 NúcleoZoulkiffiPas encore d'évaluation
- Nucleo CelularDocument15 pagesNucleo CelularVane Pons100% (3)
- Estructurasyorganelosdelacelula 10 Bio PDFDocument7 pagesEstructurasyorganelosdelacelula 10 Bio PDFPablo HernàndezPas encore d'évaluation
- Celula Animal Vs Celula VegetalDocument24 pagesCelula Animal Vs Celula VegetalVictor Flores de JezuzPas encore d'évaluation
- Nucleo CelularDocument8 pagesNucleo CelularJHARIM NOELY AUCCAPIÑA GUILLENPas encore d'évaluation
- N° 03 CelulaDocument76 pagesN° 03 CelulaYHUMY CARRION UTANIPas encore d'évaluation
- Exposición Sobre El Nucleo de La Celula EucariotaDocument14 pagesExposición Sobre El Nucleo de La Celula Eucariotajohan laraPas encore d'évaluation
- Atlas Final 2020 Grupo No.12Document48 pagesAtlas Final 2020 Grupo No.12graciela del rosario GalindoPas encore d'évaluation
- Ramón CarrilloDocument5 pagesRamón CarrilloMatías BacherPas encore d'évaluation
- Contqmiente AdmosféricoDocument1 pageContqmiente AdmosféricoMatías BacherPas encore d'évaluation
- Modelo Actual deDocument1 pageModelo Actual deMatías BacherPas encore d'évaluation
- Trabajo Práctico Del Seminario de Historia Económica Argentina y Latinoamerican ADocument4 pagesTrabajo Práctico Del Seminario de Historia Económica Argentina y Latinoamerican AMatías BacherPas encore d'évaluation
- Trabajo Práctico Del Seminario de Historia Económica Argentina y Latinoamerican ADocument3 pagesTrabajo Práctico Del Seminario de Historia Económica Argentina y Latinoamerican AMatías BacherPas encore d'évaluation
- Protocolode Detección y Actuación en Dislexia. Primer Ciclo ESO PDFDocument23 pagesProtocolode Detección y Actuación en Dislexia. Primer Ciclo ESO PDFLoli Peinado CastilloPas encore d'évaluation
- Trabajo Práctico Del Seminario de Historia Económica Argentina y Latinoamerican ADocument6 pagesTrabajo Práctico Del Seminario de Historia Económica Argentina y Latinoamerican AMatías BacherPas encore d'évaluation
- Trabajo Del Seminario de Historia Económica Argentina y Latinoamerican ADocument6 pagesTrabajo Del Seminario de Historia Económica Argentina y Latinoamerican AMatías BacherPas encore d'évaluation
- Teorias CurricularesDocument1 pageTeorias CurricularesMatías BacherPas encore d'évaluation
- AplicaciónDocument1 pageAplicaciónMatías BacherPas encore d'évaluation
- 1Document5 pages1Matías BacherPas encore d'évaluation
- Foto Toxic I DadDocument5 pagesFoto Toxic I DadMatías BacherPas encore d'évaluation
- Bacher M - Enseñanza de La CsDocument1 pageBacher M - Enseñanza de La CsMatías BacherPas encore d'évaluation
- Dialnet PsicologiaEducacionalEscolar 3642863Document25 pagesDialnet PsicologiaEducacionalEscolar 3642863Matías BacherPas encore d'évaluation
- Instituciones Edu Trabajo 1 LeandroDocument6 pagesInstituciones Edu Trabajo 1 LeandroMatías BacherPas encore d'évaluation
- 50 - HERBIFEN SUPER (2.4 D Ester 100%) - Hoja Seguridad ProductoDocument5 pages50 - HERBIFEN SUPER (2.4 D Ester 100%) - Hoja Seguridad ProductoMatías BacherPas encore d'évaluation
- Electronic A 24 Capitulos El Mundo de LaDocument399 pagesElectronic A 24 Capitulos El Mundo de LaFernando LopezPas encore d'évaluation
- Definición de ErosiónDocument5 pagesDefinición de ErosiónMatías BacherPas encore d'évaluation
- Sistemática Vegetal o Botánica VegetalDocument4 pagesSistemática Vegetal o Botánica VegetalEstefany Ramirez100% (1)
- Nueva Investigación Hilda MateusDocument181 pagesNueva Investigación Hilda MateusGreixiPas encore d'évaluation
- Morfologia de Los MicrorganismosDocument18 pagesMorfologia de Los MicrorganismosKenner Abrahan Da Rocha LabradorPas encore d'évaluation
- Clases Micro AshDocument5 pagesClases Micro AshValentina ArangoPas encore d'évaluation
- Cancer de OvarioDocument13 pagesCancer de OvarioJairo Antonio PerezPas encore d'évaluation
- Sistemas BiológicosDocument15 pagesSistemas BiológicosItzel RosasPas encore d'évaluation
- Desarrollo de MedicamentosDocument36 pagesDesarrollo de MedicamentosKurosaki_ ElizPas encore d'évaluation
- Procesamiento Postranscripcional Del ArnDocument26 pagesProcesamiento Postranscripcional Del ArnJorge Carlos Vazquez Sanchez100% (1)
- Taller de Profundización de Genética Parte 1Document7 pagesTaller de Profundización de Genética Parte 1José Nicolás Díaz VelásquezPas encore d'évaluation
- Tarea 5-EsaviDocument6 pagesTarea 5-EsavisindyPas encore d'évaluation
- Proyecto Internacional de HAPMAPDocument6 pagesProyecto Internacional de HAPMAParmando92110% (1)
- Articulo 1Document9 pagesArticulo 1Bioetica AgroindustrialPas encore d'évaluation
- Señalización celular y transducción de señales: comunicación entre las célulasDocument85 pagesSeñalización celular y transducción de señales: comunicación entre las célulasRooKyAxesoPas encore d'évaluation
- Inf.3, GC3Document5 pagesInf.3, GC3dina luz montejp riconPas encore d'évaluation
- GUIA 4 6° Grado Transporte CelularDocument2 pagesGUIA 4 6° Grado Transporte CelularGIESCYPas encore d'évaluation
- CItoDocument15 pagesCItoRaúl David Reyes LeónPas encore d'évaluation
- Esquema de Vacunación IDocument1 pageEsquema de Vacunación IKeyth Aquino NuñezPas encore d'évaluation
- La Selección NaturalDocument2 pagesLa Selección NaturalPEDRO MANUEL SOTO GUERRERO0% (3)
- Ácidos nucleicos: ADN y ARNDocument5 pagesÁcidos nucleicos: ADN y ARNwalter parraPas encore d'évaluation
- Biologia Los Tejidos - Juan Diego Mujica y Sergio Guevara 11CDocument3 pagesBiologia Los Tejidos - Juan Diego Mujica y Sergio Guevara 11CJuan Diego MujicaPas encore d'évaluation
- LacteosDocument3 pagesLacteosJhosue DavidPas encore d'évaluation
- Adn Secion de AprendizajeDocument6 pagesAdn Secion de AprendizajeRony Gonzaga Bustamante Rco100% (2)
- Clase 1. Constituyentes Del NucleoDocument39 pagesClase 1. Constituyentes Del NucleoDaniela CancinoPas encore d'évaluation
- Tallerleyesdemendelno1genticamarzo3 2014 140504073038 Phpapp02Document2 pagesTallerleyesdemendelno1genticamarzo3 2014 140504073038 Phpapp02Andres A. Rocha UsugaPas encore d'évaluation
- Directrice S Diagnostico RabiaDocument35 pagesDirectrice S Diagnostico RabiaYordy Pabel GanazhapaPas encore d'évaluation
- BIOELECTRICIDADDocument3 pagesBIOELECTRICIDADKarid Nikol RodriguezPas encore d'évaluation
- ObjetivosClonaciónGenéticaDocument4 pagesObjetivosClonaciónGenéticaRivas NairuskaPas encore d'évaluation
- Guía La Farmacia NaturalezaDocument3 pagesGuía La Farmacia NaturalezaKarenPas encore d'évaluation
- Clase 1 DR ARCOSDocument30 pagesClase 1 DR ARCOSalexis yupanquiPas encore d'évaluation
- Mutaciones causan PKU y sus efectosDocument4 pagesMutaciones causan PKU y sus efectosAylin RendonPas encore d'évaluation