Vous aimerez peut-être aussi
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (895)
- Learners ' Health and Safety Practices and Their Academic PerformanceDocument10 pagesLearners ' Health and Safety Practices and Their Academic PerformanceHira SaddozaiPas encore d'évaluation
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- Passive ROMDocument3 pagesPassive ROMCzarina FayePas encore d'évaluation
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- Industrial Attachment ReportDocument34 pagesIndustrial Attachment ReportOtsile Charisma Otsile Saq100% (1)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5794)
- Insurance CodeDocument18 pagesInsurance CodeKenneth Holasca100% (1)
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- Monitor Zoncare - PM-8000 ServicemanualDocument83 pagesMonitor Zoncare - PM-8000 Servicemanualwilmer100% (1)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (266)
- Information Technology Solutions: ADMET Testing SystemsDocument2 pagesInformation Technology Solutions: ADMET Testing Systemskrishgen biosystemsPas encore d'évaluation
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (400)
- Coatings and Waterproofing: KhalafDocument19 pagesCoatings and Waterproofing: KhalafmohamedPas encore d'évaluation
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- 07 Post Task Theory Matrix Table RevisedDocument8 pages07 Post Task Theory Matrix Table RevisedHemalyn Bereber100% (1)
- Celgene V Actavis AbraxaneDocument131 pagesCelgene V Actavis AbraxaneiphawkPas encore d'évaluation
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- JIDMR SCOPUS Ke 4 Anwar MallongiDocument4 pagesJIDMR SCOPUS Ke 4 Anwar Mallongiadhe yuniarPas encore d'évaluation
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (588)
- RA ELECTRONICSTECH CEBU Apr2019 PDFDocument12 pagesRA ELECTRONICSTECH CEBU Apr2019 PDFPhilBoardResultsPas encore d'évaluation
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
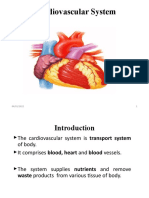
- Cardiovascular SystemDocument47 pagesCardiovascular SystemAtnasiya GadisaPas encore d'évaluation
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- Gulayan Sa Barangay OrdinanceDocument2 pagesGulayan Sa Barangay OrdinancekonsinoyePas encore d'évaluation
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (74)
- C 08 S 09Document8 pagesC 08 S 09Marnel Roy MayorPas encore d'évaluation
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (345)
- DefibrillatorDocument2 pagesDefibrillatorVasanth VasanthPas encore d'évaluation
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- Vulnerability of The Urban EnvironmentDocument11 pagesVulnerability of The Urban EnvironmentKin LeePas encore d'évaluation
- Interventional Cardiology and SurgeryDocument19 pagesInterventional Cardiology and SurgeryDEV NANDHINI RPas encore d'évaluation
- Structral DatasheetDocument254 pagesStructral DatasheetdeepakPas encore d'évaluation
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- PLAB 2 VISA Guidelines by Omar AlamDocument18 pagesPLAB 2 VISA Guidelines by Omar Alamrafew19Pas encore d'évaluation
- CA500Document3 pagesCA500Muhammad HussainPas encore d'évaluation
- Unit Plan Adult Health Nursing Unit IIDocument7 pagesUnit Plan Adult Health Nursing Unit IIDelphy VarghesePas encore d'évaluation
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2259)
- What Is A Solar Storm?Document2 pagesWhat Is A Solar Storm?Shawn SriramPas encore d'évaluation
- Meyer-Andersen - Buddhism and Death Brain Centered CriteriaDocument25 pagesMeyer-Andersen - Buddhism and Death Brain Centered Criteriautube forPas encore d'évaluation
- API Filter Press - Test ProcedureDocument8 pagesAPI Filter Press - Test ProcedureLONG LASTPas encore d'évaluation
- Marantz - dv-4200 DVD Player PDFDocument60 pagesMarantz - dv-4200 DVD Player PDFH.V KayaPas encore d'évaluation
- Oseco Elfab BioPharmaceuticalDocument3 pagesOseco Elfab BioPharmaceuticalAdverPas encore d'évaluation
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (121)
- Complete Denture TechniquesDocument6 pagesComplete Denture TechniquesJohn Hyunuk ChoPas encore d'évaluation
- Aplication Pipe and Tube - Nippon SteelDocument29 pagesAplication Pipe and Tube - Nippon Steelmatheus david100% (1)
- CHAPTER 3 Formwork Part 1Document39 pagesCHAPTER 3 Formwork Part 1nasPas encore d'évaluation
- Industrial SpecialtiesDocument103 pagesIndustrial SpecialtiesRahul ThekkiniakathPas encore d'évaluation
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)