Vous aimerez peut-être aussi
- Rapid Regioselective Oligomerization of L Glutamic Acid Diethyl Ester Catalyzed by PapainDocument7 pagesRapid Regioselective Oligomerization of L Glutamic Acid Diethyl Ester Catalyzed by PapainIvan SuarezPas encore d'évaluation
- Mutation of An Inosine Producing Strain of Bacillus Subtilis For Guanosine ProductionDocument5 pagesMutation of An Inosine Producing Strain of Bacillus Subtilis For Guanosine Production김영우Pas encore d'évaluation
- Jamur Tiram 1Document6 pagesJamur Tiram 1Muhammad AkbarPas encore d'évaluation
- Ts-2 Production of 5-Aminolevulinic Acid by Propionibacterium Acidipropionici Tistr442Document4 pagesTs-2 Production of 5-Aminolevulinic Acid by Propionibacterium Acidipropionici Tistr442Giang Huy DiệmPas encore d'évaluation
- tmp10D8 TMPDocument7 pagestmp10D8 TMPFrontiersPas encore d'évaluation
- PARTITIONING PAL ENZYMEDocument8 pagesPARTITIONING PAL ENZYMEDrVishwanatha HNPas encore d'évaluation
- Factors Affecting The Activity of Pyruvate Kinase of Acetobacter XylinumDocument6 pagesFactors Affecting The Activity of Pyruvate Kinase of Acetobacter XylinumHalima AdenPas encore d'évaluation
- 10 1007@bf00522476Document12 pages10 1007@bf00522476thiagoPas encore d'évaluation
- Lovastatin Production by Pleurotus ostreatus Affected by C:N RatioDocument4 pagesLovastatin Production by Pleurotus ostreatus Affected by C:N RatioAhmed BadrPas encore d'évaluation
- The Magic Technique For VX-680.20140730.142912Document2 pagesThe Magic Technique For VX-680.20140730.142912appeal4bottomPas encore d'évaluation
- FDADocument7 pagesFDASachithPas encore d'évaluation
- Inhibarea GlutationuluiDocument4 pagesInhibarea GlutationuluiRama KrisnaPas encore d'évaluation
- Características Del Aceite Extraído Con Fosfatasa Recombinante LIS KATERINEDocument6 pagesCaracterísticas Del Aceite Extraído Con Fosfatasa Recombinante LIS KATERINEYury MartinezPas encore d'évaluation
- SERUM GST AND EFFECT Cl4C IN RATDocument9 pagesSERUM GST AND EFFECT Cl4C IN RATToonny JasshPas encore d'évaluation
- Bte 204 Assignment: Name: Sadman Rashid Abir ID: 20136004 SEC: 01Document7 pagesBte 204 Assignment: Name: Sadman Rashid Abir ID: 20136004 SEC: 01Sadman RashidPas encore d'évaluation
- Aiba 1968 Kinetics of Product Inhibition in Alcohol FermentationDocument20 pagesAiba 1968 Kinetics of Product Inhibition in Alcohol Fermentationrishu2525Pas encore d'évaluation
- Enzymatic HydrolysisDocument8 pagesEnzymatic HydrolysisPrakash VaghasiyaPas encore d'évaluation
- Banana Polyphenoloxidase. Preparation and 2: PropertiesDocument6 pagesBanana Polyphenoloxidase. Preparation and 2: PropertiesYanely CanalesPas encore d'évaluation
- Ntiana Manshurica PDFDocument7 pagesNtiana Manshurica PDFcoffey-geoPas encore d'évaluation
- Plant Physiol.-1980-Poulton-171-5Document5 pagesPlant Physiol.-1980-Poulton-171-5Patrisia HallaPas encore d'évaluation
- Mechanism of Cytoplasmatic PH Regulation in Hypoxic Maize Root Tips and Its Role in Survival Under HypoxiaDocument5 pagesMechanism of Cytoplasmatic PH Regulation in Hypoxic Maize Root Tips and Its Role in Survival Under HypoxiaGabriel ZahariaPas encore d'évaluation
- Effect of Yeast Extract On Alpha-Amylase Synthesis by Bacillus AmyloliquefaciensDocument6 pagesEffect of Yeast Extract On Alpha-Amylase Synthesis by Bacillus AmyloliquefaciensAndalib TabassumPas encore d'évaluation
- Biochemical Studies Cocoa Bean Polyphenol Oxidase: Pat LeeDocument10 pagesBiochemical Studies Cocoa Bean Polyphenol Oxidase: Pat LeeAntra OjhaPas encore d'évaluation
- Hildegarde Esther Allen: City of Medical AND Division OF OFDocument7 pagesHildegarde Esther Allen: City of Medical AND Division OF OFJulien Patrick CebrianPas encore d'évaluation
- Insider Secrets For Vorinostat.20140729.141010Document2 pagesInsider Secrets For Vorinostat.20140729.141010plain70neckPas encore d'évaluation
- Serine Subtilis Strain Marburg: Intracellular Proteinase of Bacillus 168Document7 pagesSerine Subtilis Strain Marburg: Intracellular Proteinase of Bacillus 168Cristian Sneider Pinzon TopalPas encore d'évaluation
- Biosynthesis of Protease From Lactobacillus Paracasei: Kinetic Analysis of Fermentation ParametersDocument5 pagesBiosynthesis of Protease From Lactobacillus Paracasei: Kinetic Analysis of Fermentation ParametersGovGovsPas encore d'évaluation
- TMP 2 AF0Document4 pagesTMP 2 AF0FrontiersPas encore d'évaluation
- Artigo 1Document6 pagesArtigo 1MoleCule WitsarutPas encore d'évaluation
- 0065 2571 (72) 90019 2Document20 pages0065 2571 (72) 90019 2Muhammad Akbar SusenoPas encore d'évaluation
- Plant Physiol. 1974 Beale 291 6Document6 pagesPlant Physiol. 1974 Beale 291 6Giang Huy DiệmPas encore d'évaluation
- Biochemistry Combined Lab ReportDocument12 pagesBiochemistry Combined Lab Reportapi-283089766Pas encore d'évaluation
- Isolation and Characterization of Three Acid Phosphatase Isoenzymes from Wheat GermDocument6 pagesIsolation and Characterization of Three Acid Phosphatase Isoenzymes from Wheat GermBarry WhitePas encore d'évaluation
- Papain, Chymotrypsin and Related Proteins-A Comparative Study of Their Beer Chill-Proofing Abilities and CharacteristicsDocument5 pagesPapain, Chymotrypsin and Related Proteins-A Comparative Study of Their Beer Chill-Proofing Abilities and CharacteristicsAna PaulaPas encore d'évaluation
- Extracellular Expression of Alkaline Phytase in Pichia Pastoris InfluenceDocument7 pagesExtracellular Expression of Alkaline Phytase in Pichia Pastoris InfluenceNatalia Andrea Gómez RavePas encore d'évaluation
- Isolation and Characterization of E-64, A New Thiol Protease InhibitorDocument7 pagesIsolation and Characterization of E-64, A New Thiol Protease InhibitorHassanPas encore d'évaluation
- Phospho-opsin Phosphatase Insight Into Protamine StimulationDocument12 pagesPhospho-opsin Phosphatase Insight Into Protamine StimulationnaPas encore d'évaluation
- Hydrolysis of Beet Pulp Polysaccharides Bu Extracts of Solid State Cultures of Penicillium CapsulatumDocument6 pagesHydrolysis of Beet Pulp Polysaccharides Bu Extracts of Solid State Cultures of Penicillium CapsulatumANTONIO CELSO R DE OLIVEIRAPas encore d'évaluation
- 10 1007@BF00390820Document8 pages10 1007@BF00390820sayyed bassir ajellehPas encore d'évaluation
- List of FiguresDocument7 pagesList of FiguresSurya PrakashPas encore d'évaluation
- Synthesis and Characterization of New Amino Acid-Schiff Bases and Studies Their Effects On The Activity of ACP, PAP and NPA Enzymes (In Vitro)Document9 pagesSynthesis and Characterization of New Amino Acid-Schiff Bases and Studies Their Effects On The Activity of ACP, PAP and NPA Enzymes (In Vitro)Pavan SrivastavaPas encore d'évaluation
- Synthesis Soybean: Heme in Root NodulesDocument4 pagesSynthesis Soybean: Heme in Root NodulesEmmilyPas encore d'évaluation
- Protein Synthesis in Avocado Fruit Tissue PDFDocument4 pagesProtein Synthesis in Avocado Fruit Tissue PDFdr.sameer sainiPas encore d'évaluation
- Succinic Acid 4Document3 pagesSuccinic Acid 4Noelraj KalkuriPas encore d'évaluation
- Jsfa.274055021020161117 32447 1hvsqq With Cover Page v2Document11 pagesJsfa.274055021020161117 32447 1hvsqq With Cover Page v2Afandynibandera AfandynibanderaPas encore d'évaluation
- In The Arginine Deiminase Regulation of Enzyme Synthesis: D Haas and V Stalon A Mercenier, J P Simon, C Vander WauvenDocument6 pagesIn The Arginine Deiminase Regulation of Enzyme Synthesis: D Haas and V Stalon A Mercenier, J P Simon, C Vander WauvenAvishekh SinhaPas encore d'évaluation
- Abstract. Fermentation Conditions Were Developed in Order To Achieve Simultaneously A HighDocument9 pagesAbstract. Fermentation Conditions Were Developed in Order To Achieve Simultaneously A HighKarina A.Pas encore d'évaluation
- Biochemistry of Hormones 2Document42 pagesBiochemistry of Hormones 2Hazel Grace BellenPas encore d'évaluation
- Vol. 16, No. 1 Jurnal Ilmu Kefarmasian Indonesia, April 2018, Hlm. 45-48 ISSN 1693-1831Document4 pagesVol. 16, No. 1 Jurnal Ilmu Kefarmasian Indonesia, April 2018, Hlm. 45-48 ISSN 1693-1831Rita RakhmaPas encore d'évaluation
- Aromatic Amino Acid MetabolismDocument11 pagesAromatic Amino Acid Metabolismsoumya palavalasaPas encore d'évaluation
- Purification and Characterization of An Intracellular 13-Glucosidase From ATCC 393Document10 pagesPurification and Characterization of An Intracellular 13-Glucosidase From ATCC 393Anastasya AmandaPas encore d'évaluation
- PosterSSCHE2014 ReactEngineeringnCatalysisPosterSessionDocument1 pagePosterSSCHE2014 ReactEngineeringnCatalysisPosterSessionDiana Noor IsmailPas encore d'évaluation
- Electrophoresis and Fractionation of Wheat GlutenDocument14 pagesElectrophoresis and Fractionation of Wheat GlutensecucaPas encore d'évaluation
- Protein Structure and Folding:: Into The Reduced Toxicity of Abrus Structure - Function Analysis and InsightsDocument21 pagesProtein Structure and Folding:: Into The Reduced Toxicity of Abrus Structure - Function Analysis and InsightsStephanie WilliamsPas encore d'évaluation
- Isolasi Protein Dan Produksi Konsentrat Protein Daun (KPD) Sebagai Suplemen Pakan TernakDocument7 pagesIsolasi Protein Dan Produksi Konsentrat Protein Daun (KPD) Sebagai Suplemen Pakan TernakSiti MufarohahPas encore d'évaluation
- Kinetic Analysis of Red Pigment and Citrinin Production by MonascusDocument7 pagesKinetic Analysis of Red Pigment and Citrinin Production by MonascusArely DlrcPas encore d'évaluation
- At Pa Sal BrevisDocument5 pagesAt Pa Sal BrevisNils Huaman CastillaPas encore d'évaluation
- Jimb 0055Document11 pagesJimb 0055thiagoPas encore d'évaluation
- Co- and Post-Translational Modifications of Therapeutic Antibodies and ProteinsD'EverandCo- and Post-Translational Modifications of Therapeutic Antibodies and ProteinsPas encore d'évaluation
- Fluid Mechanics, Bernoulli's Principle and Equation of ContinuityDocument10 pagesFluid Mechanics, Bernoulli's Principle and Equation of ContinuitySheerin SulthanaPas encore d'évaluation
- Unesco - Eolss Sample Chapters: Lysine Biosynthesis in Bacteria - An Unchartered Pathway For Novel Antibiotic DesignDocument11 pagesUnesco - Eolss Sample Chapters: Lysine Biosynthesis in Bacteria - An Unchartered Pathway For Novel Antibiotic DesignSheerin SulthanaPas encore d'évaluation
- BCH440 (8) 32-33Document20 pagesBCH440 (8) 32-33Sheerin SulthanaPas encore d'évaluation
- Chapter 26 Phylogeny and The Tree of LifeDocument41 pagesChapter 26 Phylogeny and The Tree of LifeSheerin Sulthana100% (1)
- Human Genome ProjectDocument57 pagesHuman Genome ProjectDhaval ParekhPas encore d'évaluation
- Appl. Microbiol. 1968 Kato 1200 6Document8 pagesAppl. Microbiol. 1968 Kato 1200 6Sheerin SulthanaPas encore d'évaluation
- A Brief Review of Plants Having Anti Cancer PropertyDocument8 pagesA Brief Review of Plants Having Anti Cancer PropertysharammaPas encore d'évaluation
- A Business-Oriented Overview of Intellectual Property For Law StudentsDocument16 pagesA Business-Oriented Overview of Intellectual Property For Law StudentsSheerin SulthanaPas encore d'évaluation
- Biological Safety Levels: Endia Ford Lori Gladney Izabella OsakweDocument34 pagesBiological Safety Levels: Endia Ford Lori Gladney Izabella OsakweSheerin Sulthana100% (1)
- 32721Document33 pages32721prasadbheemPas encore d'évaluation
- Trade-Related Intellectual Property Rights (Trips)Document20 pagesTrade-Related Intellectual Property Rights (Trips)Sheerin SulthanaPas encore d'évaluation
- Elements of Biosafety: Janet Peterson Jpeterso@accmail - Umd.edu 405-3975Document28 pagesElements of Biosafety: Janet Peterson Jpeterso@accmail - Umd.edu 405-3975Sheerin SulthanaPas encore d'évaluation
- Bioethics Timeline and ControversiesDocument18 pagesBioethics Timeline and ControversiesSheerin SulthanaPas encore d'évaluation
- Trade-Related Intellectual Property Rights (Trips)Document20 pagesTrade-Related Intellectual Property Rights (Trips)Sheerin SulthanaPas encore d'évaluation
- Analysis On The Impact of Madrid Protocol For The Economies of Developing CountriesDocument62 pagesAnalysis On The Impact of Madrid Protocol For The Economies of Developing CountriesSheerin Sulthana0% (1)
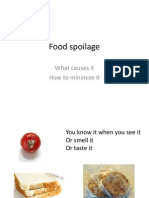
- Foodborne IllnessDocument13 pagesFoodborne IllnessSheerin SulthanaPas encore d'évaluation
- Internet and NetworkingDocument42 pagesInternet and Networkingآصف رضاPas encore d'évaluation
- Radioimmunoassay: Advanced Research Techniques in Basic Medical SciencesDocument23 pagesRadioimmunoassay: Advanced Research Techniques in Basic Medical SciencesKiran FriendooPas encore d'évaluation
- Biological DatabaseDocument19 pagesBiological DatabaseMahesh Yadav100% (8)
- ATCCDocument3 pagesATCCSheerin SulthanaPas encore d'évaluation
- Dimensions of Product Quality ManagementDocument5 pagesDimensions of Product Quality ManagementSheerin SulthanaPas encore d'évaluation
- (WWW - Entrance Exam - Net) Animal BiotechDocument34 pages(WWW - Entrance Exam - Net) Animal BiotechSheerin SulthanaPas encore d'évaluation
- The Range of Quality: Quality of Design Quality of Materials Quality of ProductsDocument4 pagesThe Range of Quality: Quality of Design Quality of Materials Quality of ProductsKin SkyePas encore d'évaluation
- Lecture 21 - Respiratory SystemDocument59 pagesLecture 21 - Respiratory SystemSheerin SulthanaPas encore d'évaluation
- Foods Poli AgeDocument23 pagesFoods Poli AgeSheerin SulthanaPas encore d'évaluation
- Emerging: Brewing Is The Production ofDocument16 pagesEmerging: Brewing Is The Production ofSheerin SulthanaPas encore d'évaluation
- Pricing ProblemsDocument2 pagesPricing ProblemsSheerin SulthanaPas encore d'évaluation
- 06iso Allo Idio09Document18 pages06iso Allo Idio09Sheerin SulthanaPas encore d'évaluation
- 10 1371-Journal Pone 0065654 g003Document1 page10 1371-Journal Pone 0065654 g003Sheerin SulthanaPas encore d'évaluation
- Order Details for Order #10105Document2 pagesOrder Details for Order #10105Mohamed HarbPas encore d'évaluation
- A Thesis 123Document77 pagesA Thesis 123Meli SafiraPas encore d'évaluation
- Closed Coke Slurry System: An Advanced Coke Handling ProcessDocument33 pagesClosed Coke Slurry System: An Advanced Coke Handling ProcessFayaz MohammedPas encore d'évaluation
- Department of Mechanical Engineering Polytechnic Sultan Haji Ahmad Shah Kuantan, Pahang DJJ 30122-CADDocument2 pagesDepartment of Mechanical Engineering Polytechnic Sultan Haji Ahmad Shah Kuantan, Pahang DJJ 30122-CADAbdul MalikPas encore d'évaluation
- Positive Role The Indian Youth Can Play in PoliticsDocument2 pagesPositive Role The Indian Youth Can Play in PoliticsArul ChamariaPas encore d'évaluation
- Basic Engineering & Site DataDocument13 pagesBasic Engineering & Site DataBalasubramanianPas encore d'évaluation
- Mouse Molecular Genetics Student Activity 2Document7 pagesMouse Molecular Genetics Student Activity 2Jonathan ZhouPas encore d'évaluation
- OHS Policies and Guidelines (TESDA CSS NC2 COC1)Document1 pageOHS Policies and Guidelines (TESDA CSS NC2 COC1)Anonymous fvY2BzPQVx100% (2)
- Risk Culture Assessment QuestionnaireDocument3 pagesRisk Culture Assessment QuestionnairemohamedPas encore d'évaluation
- Philadelphia University Faculty of Engineering and Technology Department of Mechanical EngineeringDocument8 pagesPhiladelphia University Faculty of Engineering and Technology Department of Mechanical EngineeringTamer JafarPas encore d'évaluation
- Dictionary of Oil Industry TerminologyDocument79 pagesDictionary of Oil Industry Terminologyniksharris100% (22)
- MSDS Chrome Catalyst Safety SheetDocument4 pagesMSDS Chrome Catalyst Safety Sheetstavros7Pas encore d'évaluation
- Muhammad Zahrandhika Bastian-3Document2 pagesMuhammad Zahrandhika Bastian-3dhika zahrandhikaPas encore d'évaluation
- Teaching Load FinalDocument12 pagesTeaching Load FinalEdward BarberPas encore d'évaluation
- Disaster Readiness Exam SpecificationsDocument2 pagesDisaster Readiness Exam SpecificationsRICHARD CORTEZPas encore d'évaluation
- MY LIFE VISION, MISSION AND CORE VALUES BMEC 2W 2122Document4 pagesMY LIFE VISION, MISSION AND CORE VALUES BMEC 2W 2122Nikolai Avery NorthPas encore d'évaluation
- C 6 Slings SafetyDocument29 pagesC 6 Slings SafetyAshraf BeramPas encore d'évaluation
- 7 Robert Boyle and Experimental Methods: © 2004 Fiona KisbyDocument8 pages7 Robert Boyle and Experimental Methods: © 2004 Fiona Kisbydaveseram1018Pas encore d'évaluation
- Guide To Preparing Tax Research MemosDocument2 pagesGuide To Preparing Tax Research MemoscglaskoPas encore d'évaluation
- Formative vs Reflective Measures of Organizational CoordinationDocument20 pagesFormative vs Reflective Measures of Organizational Coordinationmasterling880% (1)
- REV 0 Material Selection Diagram MSD Blueprint IncDocument5 pagesREV 0 Material Selection Diagram MSD Blueprint IncJohn Patrick Sanay NunezPas encore d'évaluation
- CS4000 Data SheetDocument6 pagesCS4000 Data SheetJuan Luis EspinosaPas encore d'évaluation
- English 9 - Q2 - M5Document16 pagesEnglish 9 - Q2 - M5myraPas encore d'évaluation
- Simulation of BJT Amplifier: Course - Section: ECE20L-E06 Group NumberDocument10 pagesSimulation of BJT Amplifier: Course - Section: ECE20L-E06 Group NumberLuch ÜPas encore d'évaluation
- Waffle Slab - WikipediaDocument15 pagesWaffle Slab - WikipediaBryan PongaoPas encore d'évaluation
- Academic SummaryDocument4 pagesAcademic SummaryJacqui PendergastPas encore d'évaluation
- Personal Data Form: World English Placement Test Package Photocopiable © 2011 Heinle, Cengage LearningDocument2 pagesPersonal Data Form: World English Placement Test Package Photocopiable © 2011 Heinle, Cengage Learningadri shimizuPas encore d'évaluation
- Unsaturated Polyester Resins: Chemistry and Technology: Piotr Penczek (U) Piotr Czub Jan PielichowskiDocument2 pagesUnsaturated Polyester Resins: Chemistry and Technology: Piotr Penczek (U) Piotr Czub Jan Pielichowskiae0011979Pas encore d'évaluation
- Radiation Safety Officers Handbook A PDFDocument100 pagesRadiation Safety Officers Handbook A PDFAlejandro Zubiate100% (1)
- Ms Word McqsDocument12 pagesMs Word McqsUraiBa AnsaRi0% (1)