Académique Documents
Professionnel Documents
Culture Documents
2075-7626-1-SM - Pdfcell Shwan PDF
Transféré par
cibert40 évaluation0% ont trouvé ce document utile (0 vote)
35 vues11 pagesTitre original
2075-7626-1-SM.pdfcell shwan.pdf
Copyright
© © All Rights Reserved
Formats disponibles
PDF, TXT ou lisez en ligne sur Scribd
Partager ce document
Partager ou intégrer le document
Avez-vous trouvé ce document utile ?
Ce contenu est-il inapproprié ?
Signaler ce documentDroits d'auteur :
© All Rights Reserved
Formats disponibles
Téléchargez comme PDF, TXT ou lisez en ligne sur Scribd
0 évaluation0% ont trouvé ce document utile (0 vote)
35 vues11 pages2075-7626-1-SM - Pdfcell Shwan PDF
Transféré par
cibert4Droits d'auteur :
© All Rights Reserved
Formats disponibles
Téléchargez comme PDF, TXT ou lisez en ligne sur Scribd
Vous êtes sur la page 1sur 11
LA CELULA DE SCHWANN
Adriana del Pilar Lopez Lornbana * y Hernn Hurtado Giraldo *
La clula de Schwann que constituye la gla del SNP, adems de ser el soporte
estructural para los axones en dicho sistema, tiene la funcin de producir la
mielina, una organela de gran importancia en los procesos de neuroconduccin.
De la integridad de esta clula dependen el desarrollo estructural y metablico del
axn, as mismo se ha reconocido desde hace varios anos el papel primordial que
juega ella, en los procesos de regeneracin del SPN posterior a una injuria, en
cuyo caso reinician la proliferacin para producir una gua de regeneracin del
nervio oerifrico. En sta revisin se contemolaran alaunos de los ountos
-
relacioiados con su origen, desarrollo, estructuia, relacin con el axon ;el tipo
de patologas que pueden alterarla; igualmente se resalta la utilidad de loscultivos
decel ul as de Schwann para el estudio de los procesos de mielinizacin,
desmielinizacin, regeneracin post-traumatica y respuesta a agentes infeccio-
sos.
INTRODUCCION ORIGEN, PROLIFERACION
Y DESARROLLO
Las clulas de Schwann, descritas desde
1839 por Theodor Schwann, constituyen la glia
del sistema nervioso perifrico (SNP). Rodean
todos los axones del nervio, en unos casos
envolviendo con su citoplasma varios de ellos y
en otros casos elaborando la vaina de mielina
alrededor de los de mayor dimetro. Estas clu-
las cumplen mltiples funciones relacionadas
con la proteccin y el soporte metablico axonal.
Adems contribuyen en los procesos de conduc-
cin nerviosa, as como en los mecanismos de
regeneracin de los axones lesionados. En este
artculo se har una revisin de los aspectos ms
relevantes de estas clulas en cuanto a su ori-
Origen
Con excepcin de ciertos ganglios craneales
sensoriales, los diferentes tipos de clulas del
SNP se derivan de una estructura embrionaria
transitoria, la cresta neural (1,2,3), aunque tam-
bin se ha discutido que un origen parcial de la
regin ventral de la cresta neural no puede ex-
cluirse para las clulas de Schwann. Se desco-
noce en que estadio embrionario las clulas de
la cresta neural comienzan a diferenciarse en
clulas de Schwann, pero se sabe que necesitan
el contacto axonal para ello (4,5).
gen, desarrollo, caractersticas morfolgicas y
proliferacin. Adems se explicar el proceso de
El desarrollo del nervio perifrico es un proce-
formacin de la mielina y la importancia de la
so ordenado. Inicialmente un grupo grande de
actividad funcional de la clula de Schwann en
axones se rodea de unas Pocas clulas de
relacin con el axn al igual que su dependencia
Schwann, las cuales se organizan de tal forma
de este ltimo para llevar a cabo sus propias
que constituyen una cubierta o tubo (6). La parte
funciones.
externa de este tubo est delineada por una
M.0 Grupo de Neurobologa del I.N.S.
** P h D Jefe del Grupo de Neurobologa del I.N.S.
AORiNA DEL PILAR LOPEZ Y HERNAN HURTADO GIRALDO
lmina basal, escencialmente producida por la perifrico como es el caso de las neurofibroma-
clulamisma, la parte internadel tubocorrespon- tosis y de los fibromas acsticos (15,37).
de al compartimiento interior de la clula de
Schwann separada del axolema unos 20-30 nm
La naturaleza Y origen de 10s factores que
(7). L~~~~ de la invasin de los axones desnudos
estan implicados en estos procesos proliferativos
por las-clulas qliales proliferantes, hay una
SOn claros an
distribucin diferencial de los axones. LOS de
dimetro mayor de 1 un- se disponen en relacin
1 :1 con las clulas de Schwann destinadas a ser
mielinizadoras. En los axones destinados a no
ser mielinizados (50-80%), la relacin es de una
clula de Schwann por 5-1 8 axones, los cuales
son rodeados por una prolongacin de la clula
glial (8). Los dos tipos de clulas de Schwann
tienen caractersticas moleculares especficas a
las que se har mencin posteriormente.
Los nervios perifricos mayores, como el nervio
citico, se pueden identificar como una discreta
estructura anatmica en un embrin de rata de
14-15 dias (9). En este momento los nervios
consisten de bandas de axones acompahados
de unas clulas de rpida proliferacin que care-
cen de lmina basal y se muestran como proce-
sos laminares aplanados (10). Estas clulas se
identifican como precursoras de las clulas de
Schwann con baseen sus propiedadesantignicas,
pero en trminos morfolgicos las clulas precur-
soras y su relacin con los axones en desarrollo
no han sido estudiadas en detalle (1 1).
Proliferacin
La proliferacin de las clulas de Schwann,
durante el desarrollo del nervio periferico es
intensa. Numerosas evidencias indican que di-
cha proliferacin es dependiente de una seiial
mitognica provista por el axn en crecimiento
an no claramente establecida (12,13,14).
Las clulas de Schwann proliferan bsica-
mente en tres contextos:
Se sabe que las clulas de Schwann pueden
proliferar en respuesta a un amplio rango de
factores solubles (cuadro 1). Estos factores se
cree ejercen su funcin al aumentar los niveles
de AMPc intracelular (15,17.). Algunas observa-
ciones hechas en cultivo sugieren que el PDGF
(factor decrecimiento derivado de plaquetas) y el
FGF (factor de crecimientofibroblstico) pueden
estimular la proliferacin de lasclulas de Schwann
si estas se exponen a condiciones ambientales
que eleven el AMP cclico intracelular.(l8). En el
caso del TGF (factor de crecimiento transforma-
dor) beta su poder mitognico es potenciado por
la aplicacin de la forscolina (1 1,16). Las res-
puestas proliferativas de las clulas de Schwann
durante la degeneracin walleriana varian con la
edad del animal. La mayor parte de este tipo de
respuestas han sido valoradas en lesiones de
nervio en animales adultos. Por otro lado, estu-
dios recientes in vivo han mostrado una relacin
inversa entre la edad del animal y la capacidad
proliferativa de sus clulas gliales, lo que parece
estar relacionado con el pobre estado de
mielinizacin de los axones (1 2). La mitosis de
las clulas de Schwann alcanza un pico mximo
al tercer da posterior a la seccin en el nervio
Cuadro 1.Molculas expresadas por clulas de Schwann
mielinizadoras y no mielinizadoras bajo condi-
ciones normales, durante el desarrolo o des-
ous de la lesin de un nervio.
Molcula Mielinizadoras No Mielinizadoras
Normales lesiones
adultas o desarrollo
1. Durante el desarrollo normal del nervio perif-
rico.(l5,16).
Galactocerebros~do
NGF
5-100
2. Luego de una lesin del nervio, por trauma
Ran~P
mecnico por neurotoxinas o enfermedades
Laminina
desmielihfzantes (16). Colageno tipo IV
Heparan Sulfato
3. En los tumores de clulas de Schwann, pue- N-CAM
den proliferar sobre cualquier tipo de nervio
GFAp
citico del ratn adulto. En nervios embrionarios
o neonatalessin embargoel ndice de marcajede
las clulas de Schwann disminuye despus de la
transeccin. Esto se ha equiparado a estudios in
vitro, en los cuales se ha encontrado que la
proliferacin de estas clulas se lleva a cabo en
aquellas relacionadas con mielina y que la pro-
liferacintermina rpidamente cuando las neuritas
jovenes son seccionadas sin que hayan comple-
tado su mielinizacin (19). Estos experimentos
se han podido comprobartambin en los cultivos
donde al remover los axones y restos de mielina
se reduce la captacin de timidina tritiada. Esta
reduccin es menor en los ratones P30 (da 30
postnatal), que en los ratones P10 (da 10 postnatal)
Se haencontradoinvitro, queclulasdeSchwann
preparadas a partir de nervio citico de rata de
seis dias postnatales proliferan ms vigorosa-
menteen respuestaafracciones enriquecidas en
mielina que aquellos de ratas de dos das de
edad (20).
Los mecanismos que tienen que ver con la
cesacin de la proliferacin en el desarrollo del
nervio normal e in vitro son oscuros an. Una
posibilidad muy interesante, es queestefenme-
no est bajo control autocrino negativo, mediante
una proteina neural antiproliferativa, un factor
protico que inhibe la sintesis de ADN en clulas
de Schwann estimuladas por mitgenos (18).
Desarrollo
El desarrollo de las clulas de Schwann, al
igual que el de muchos otros sistemas, se carac-
teriza por unafase embrionaria y unaneonatal de
rpida proliferacin seguidas por la cesacin de
la proliferacion y su diferenciacin final (1 1,21).
En su desarrollo normal hay dos etapas impor-
tantes: una migratoria y la otra mielinizadora. Las
clulas de Schwann en su fase migratoria son
largas, bipolares, con una composicin rica en
microfilamentos y carecen de Imina basal y
mielina (22). Se disponen en el nervio, sobre los
axones en su posicin final dividiendolo en pe-
quetias familias de varios axones rodeados de
una o dos clulas de Schwann. Estas continuan
proliferando y el ririiero de axories por clula de
Schwann disminuye.(4,8). Sirnultaneamente los
axones ms grandes (mayores de 1 urri) erripie-
zan asegregarse de sus similares y a aislarse en
el citoplasma de una sola clula de Schwann. En
este estadio los espacios de tejido conectivo en
el n e ~ i o ya se han desarrollado mejor y las
clulas de Schwann estn ya ensamblando Imi-
na basal.(l4). Este es un evento muy importante
puesto que la futura maduracin de las clulas y
en particular la mielinizacin se cree que depen-
den del correcto ensamblaje de dicha Imina
(23). En el nervio citico de rata recin nacida las
clulas se encuentran ya en relacin 1 :1 con los
axones y las primeras envolturas de mielina se
forman durante las primeras 24 horas de vida. En
esta especie la maduracin de las clulas de
Schwann no formadoras de mielina ocurre du-
rante la tercera semana postnatal y su desarrollo
es completo 2-3 semanas ms tarde, lo cual
coincide con lacesacin de la proliferacin de las
clulas de Schwann (1 1).
MORFOLOGIA Y ULTRAESTRUCTURA
DE LA CELULA DE SCHWANN
En el nervio normal la clula de Schwann est
rodeada de una Imina basal de 20-30 nm de
espesor. Esta Imina es continua an a nivel de
los nodos de Ranvier (24). El citoplasma de las
clulas de Schwann es rico en organelas. El
aparato de Golgi se localiza cerca al ncleo y las
cisternas de retculo endoplsmico rugoso se
encuentran dispersas en toda la clula; tambin
pueden observarse abundantes lisosomas, cuer-
pos multivesiculares, grnulos lipdicos y de
glicgeno (4). La membrana plamtica muestra
vesculas pinocitticas. Se hallan mitocondrias
pequeas y redondeadas a travs del soma. El
ncleoesaplanadoy seorientalongitudinalmente
a lo largo de la fibra, y su heterocromatina se
distribuye perifricamente (4,25).
Se reconocen cuatro fenotipos de clulas de
Schwann sobre bases morfolgicas:
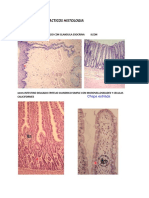
1. Clulas de Schwann formadoras de mielina,
que envuelven solamente axones de ms de
1 um de dimetro (figura 1 .)
2. Clulas de Schwann no mielinizantes, que
rodean mltiples axones pequeos (figura 1 .).
3. Clulas satlites que rodean los pericariones
neuronales en los ganglios (figura 2.).
AORINA DEL PILAR LOPEZ Y HERNAN HURTADO GIRALOO
Flgura 1. Corte transversal de nervio perifrico de rata
adulta que ilustra fibras mielinizadas (M) y no mi eni zadas
(U). Un fibroblasto (flecha) emite largas prolongaciones en
la matriz extracelular que contene colgeno (cf). 4.500 X.
Figura 2. Ganglio espina1 (ganglio de a raiz dorsal) de rata
adulta. Se ilustran neuronas sensoriales (flechas) v ceiulas
sateites (cabeza de flecha). 600 X.
21
4. Agregados tubulares de clulas de Schwann
(Bandasde Bngner) en lossegmentosdistales
de los nervios seccionados (8).
Estos fenotipos de clulas de Schwann son
neuronalmente determinados y parecen ser
interconvertibles. El fenotipo de las clulas de
Schwann durante la diferenciacin es fuerte-
mente regulado por contacto axonal; en estudios
in vitro el fenotipo de estas clulas puede ser
convertido de no mielinizante a mielinizante sim-
plemente forzando una interaccin con axones
que son normalmente mielinizados. Esta plasti-
cidad es notoria si se tienen en cuenta las dife-
rencias bioqumicas y morfolgicas de lasclulas
mielinizadoras y no mielinizadoras maduras (26)
En la clulade Schwann se han descrito cinco
dominios citoplasmticos que permiten estudiar
la organizacin metablica de la clula mieliniza-
dora (figura 3.). Son ellos:
1. Dominio perinuclear.
2. Dominio de canales citoplasmticos superfi-
ciales.
3. Cisuras de Schmidt-Lanterman.
DP: Dominio perinuciear
N: Nucleo
A: Axin
DPN: Dominio perinodal (Asas paranodales)
DA: Dominio adavonal
CSL Cisuras de Sehmidth Lanterman
Fi gura3 Dom~nioscitoplasmiticosdeiaceluladeSchwann,
Organizacon metablica.
LA CELULA DE SCHWANN
4. Asas paranodales.
5. Dominio adaxonal
El eje de la actividad metablica de la clula
de Schwann parece encontrarse en el dominio
perinuclear, el cual contiene la mayora de
organelas implicadas en la sntesis de Ipidos y
proteinas (25). Las protenas bsicasde lamielina
y losfosfolpidossintetizadosenel reaperinuclear
viajan a lo largo de los canales citoplsmatico$ a
sus lugares o sitios de integracin en las capas
de mielina compacta. En el dominio perinuclear
es donde se sintetizan predominantemente las
protenas de la mielina. No ha sido posible en-
contrar lipidos o protenas formados reciente-
mente en las asas paranodales o en las cisuras
de Schmidt-Lanterman, lo cual ha llevado a con-
cluir que estos dominios no participan en el
movimiento de estas molculas hacia las capas
compactas de mielina.
INTERACCION CELULA
DE SCHWANN-AXON
Las clulas de Schwann se definen usual-
mente por su cercana disposicin con las fibras
nerviosas. Se sabe ahora que hay mltiples
interacciones funcionales y bioquimicas entre la
clula de Schwann y el axn (1). Los axones
tienen un papel dominante al influir en el nmero
de clulas de Schwann, l a entrada de ellas en el
ciclo celular, su supervivencia, la expresin de
una variedad de marcadores fenotpicos, su pro-
duccin de Imina basal y de mielina y el mante-
nimiento de sta ltima (1,7,10). Estas clulas
son responsables de la generacin de su Imina
basal, secretan al menos tres de sus principales
Bajo nivel de m a
RNAm y colgeno
tipo IV en el medio
incremento 10
veces en el
nivel de RNAm
laminina y wlgeno tipo IV
Figura4. RegulaciOndelaactividaddelaciuladeSchwann
por el axn.
Mediante estudios inmunohistolgicos se ha
demostrado que las clulas de Schwann expre-
san molculasdesuperficiey protenasdemielina
solo si estn en presencia de axones (2,30,31).
Esto ha sido comprobado in vitro deprivando las
clulas del contacto neurona1 con lo cual se
detiene el proceso de expresin de dichas mol-
culas (7,29). Igualmente ocurre con la expresin
del galactocerebrsido C (Gal C) sobre su super-
ficie (29)
En general las consecuencias mejor docu-
mentadas de la relacin clulade Schwann-axn
son la induccin de la proliferacin de estas
clulas y la formacin de mielina. Se ha prestado
menos atencin a la modulacin de las propieda-
des axonales por la gla, aparte de la conocida
funcin de permitir la "conduccin saltatorial' del
potencial de accin (1 6). Recientementealgunos
estudios han mostradoquelasclulasdeSchwann
v la mielinizacin ~eri fri ca ~ueden modular di-
componentes como son: la laminina, el colgeno
iectamente la estrictura y funcin axonal. Se ha
tipolVyelproteoglicano heparnsulfato(27,28,29).
observado que la desmielinizacin disminuye la
Para que stas funciones ocurran, es necesaria
la presencia de neuronas, bsicamente para el
ensamblaje y organizacin de dicha Imina basal
(23). Se cree que una forma como las neuronas
regulan la formacin de la Imina basal por las
clulas de Schwann es mediante el incremento
de una seal difusible que acta estimulando la
rata de transcripcin de ciertos genes que codi-
fican para componentes de la Imina basal (figu-
ra 4) (27)
fosforilacin de neurofilamentos axonales, el di-
metro axonal y el transporte axonal lento. incre-
mentando significativamente la densidad de los
neurofilamentos (32).
Algunos trabajos utilizando anticuerpos es-
pecficos para canales de sodio han mostrado
que el contacto entre clula de Schwann y axn
es esencial en la regulacin de la distribucin de
canales de sodio sobre la membrana axonal
ADRNA DEL PILAR LOPEZ Y HERNAN HURTADO GIRALOO
(33,34). Con los cambios que ocurren durante la
mielinizacin estos canales empiezan a concen-
trarse en el axolema a nivel nodal mientras que
los canales de potasio se acumulan en las regio-
nes paranodal e internodal (30,34).
MIELINA Y MlELlNlZAClON
La mielina es una organela celular nica, una
extensin modificadade lamembranaplasmtica
de la clula de Schwann en el nervio perifrico
(35). Su elaboracin depende de la expresin
regulada de un conjunto de genes en la gla
mielinizante, para lo cual las clulas de Schwann
requieren del contacto axonal, al igual que para
mantener la expresin de dichos genes (36). La
mielina acta como un aislante elctrico que
facilitalaconduccindel impulso nervioso (16,35)
En los axones amielnicos, el impulso se transmi-
te por circuitos locales de corrientes inicas (26).
Esto hace que la conduccin sea ms lenta si se
compara con la llamada "conduccin saltatoria"
de las membranas de los axones mielinizados,
en las cuales la excitacin ocurre a nivel de los
nodos de Ranvier (regiones del axn entre dos
segmentos de mielina). En estas regiones, en las
cuales no hay cubierta mielnica y el axn esta
rodeado solamente de lmina basal (34), el im-
pulso local generado all no puede fluir por la alta
resistencia de la vaina y por lo tanto salta y
despolariza la membrana a nivel del siguiente
nodo (26)
En la ltima dcada se han realizado mlti-
ples investigaciones, encaminadas a encontrar y
entender cmo se elaboran, ensamblan y orga-
nizan los elementos de la mielina en una vaina
compacta. El mecanismo de depsito de mielina
es oscuro an. Durante la mielinizacin hay un
incremento de la longitud internodal, en el dime-
tro axonal y en el nmero de capas de mielina,
pero el momento en que se detiene su depsito
an no se ha establecido (36). Hay al menos dos
na plasmtica de las clulas. Esta ltima hipte-
sis es la ms aceptada hasta el momento (37).
La mielina in situ tiene un contenido de agua
de casi el 40%. Su masa seca se caracteriza por
una alta proporcin de lpidos (70.85%) y por lo
tanto baja proporcin de protenas en contraste
con la mayora de membranas que tiene esta
proporcin invertida (26).
Las protenas de mielina en el SNP son dife-
rentes de las del SNC, en variedad y proporcin
(cuadro 2). Los nervios perifricos no contienen
el proteolpidoproteico caractersticodelamielina
central y su contenido en MBP (Protenas bsi-
cas de la mielina) es bajo. Una sola protena, la
PO (P zero), constituye ms del 50% de las
protenas de mielina del SNP (35). PO es una
glicoprotena fosforilada, sulfatada y acilada de
28 kd. Es el principal componente estructural de
la mielina perifrica, expresada exclusivamente
por clulas de Schwann mielinizantes. Se ha
propuesto que su funcin es la contrapartida
perifrica de la funcin central que tiene el
proteolpido proteico, es decir promover laforma-
cin y laestabilizacinde laestructuramultilamelar
compacta de la mielina (30). PO y MBP se loca-
lizan exclusivamente en la regin compacta de la
mielinaen las clulasdeSchwann mielinizadoras
(9). Estas protenas no son detectadas antes de
la formacin de mielina, mientras el Gal-C
(Galactocerebrsido C) y la MAG (Glicoproteina
Asociada a la Mielina) aparecen antes de que se
inicie este fenmeno y su porcentaje de expre-
sin disminuyedramticamenteunavezse inicia
la mielinizacin (8,36).
Cuadro 2 .Mitogenos para clulas de Schwann in vitro
Toxina del clera
Factor de maduracin glial
Factor de crecimiento fibroblstico
puntos de vista de cmo las clulas de Schwann
de crecimiento derivado de las plaqueias
forman susvainas de mielinaenformade espiral.
Laminina
Segn el primero, el cuerpo de las clulas rota
alrededor del axn, mientras aue el seaundo
Fibronec"na
sostiene que la mielina es formada por un creci.
Factor de crecimiento derivado de Schwanomas
miento continuo de lacapa internade la membra- Forscoina 1
LA CELULP DE SCHWANN
El contenido de MBP vara del 5.18% de las
protenas totales en el SNP, en contraste con un
20.40% en el SNC. Son proteinas pequehas (1 4-
21 kd) que se han localizado inmunocito-
qumicamente en las lneas densas de la mielina.
Las MBP incluyen cuatro protenas, siendo las
ms prominentes P, (1 8 kD) y especialmente P,
(14 kD) que es aproximadamente el 1% en roe-
dores y el 5% en humanos (4, 26). Esta ltima
considerada clsicamente como exclusiva del
SNP, se ha encontrado en pequehas cantidades
en el SNC de algunas especies. La P, es el
antgeno para la neuritis alrgica experimental,
la contraparte en el SNP de la encefalitis alrgica
experimental (26).
El contenido de protenas de alto peso mole-
cular es menor en el SNP si se compara con el
SNC. La MAG se expresa en niveles relativa-
mente bajos en el SNC y en el SNP. Esta mol-
cula comparte algunas caractersticas con las
proteinas de adhesin N-CAM y L-1 (14,26,38).
Mediante estudios inmunohistoqumicos se ha
revelado que la MAG tiene una localizacin
subcelular en la clulas mielinizantes y est
aparentemente excluida de la mielina compacta,
expresndose principalmente en el margen
periaxonal de la vaina de mielina, en las cisuras
de Schmidt-Lanterman y en lasasas paranodales
(35). Ella puede ser detectada en estados muy
tempranos de mielinizacin. Estas observacio-
nes han llevado a formular la hiptesis de que la
MAG juega un papel en la mediacin de los
eventos de reconocimiento axn-gla que prece-
den a la mielinizacin, dando una seal inicial
para el depsito de mielina (38). Es interesante
que su estructura al igual que la de la PO se
relaciona con la de las inmunoglobulinas. La
MAG contiene la secuencia Arg-Gly-Asp, un
tripptido que media la adhesin de muchas
molculas de matriz extracelular a sus recepto-
res. Esta caracterstica se ha interpretado como
una evidencia indirecta que sostiene la hiptesis
de que la MAG es una molcula de adhesin
importante parael iniciode la mielinizacin (26,38).
cuantitativas. El SNPtiene menoscerebrsidos
y sulftidos y ms esfingomielina que el SNC
(4).
En cuanto a las clulas de Schwann no
formadoras de mielina, se sabe que sobrepasan
en nmero a las mielinizadoras, constituyendo
una importante categora de gla perifrica. Se
sabe pocoacercadelascaractersticas moleculares
de estas clulas, pero se conoce que expresan
algunas proteinas que no se detectan en clulas
mielinizadoras, pero s en astrocitos y en clulas
de la gla entrica (Cuadro 3. 11,39). Las clulas
deschwann no mielinizadoras expresan IaGFAP
(Proteina glial fibrilar cida) que inicialmente se
consideraba como especfica de los filamentos
intermediariosdeastrocitos, y otras tresproteinas
desuperficiedenominadas Ran-2, antgenoA5E5
y laN-CAM (molculadeadhesin neural) (31,39)
La molcula de adhesin L- l es detectable tanto
en clulasmielinizadorascomo no mielinizadoras,
pero en cantidades diferentes dependiendo de
su fase de desarrollo pues su expresin disminu-
ye notablemente una vez inician la fase
mielinizadora. Las clulas no mielinizadoras
permanecen positivas para L-1 y N-CAM an en
la vida adulta (39,40).
Cuadro 3. Comparacin de proteinas de mielina en SNC y
SNP en adulto
Proteina SNC SNP
P2 Trazas r) 0.05-0,1%
PO >so %
MAG - 0.1 46
MBP 30 % 5-18%
PLP 50 % -
MAG:Glicoproteina asociada a la mielina.
MBP: Proteina bsica de i a mieiina.
PLP: Proteoiipido proteico o proteina de Folch-Lees
SNC: Sistema Nervioso Central
SNP: Sistema Nervioso Perifrico
* , P2 clsicamente considerada exclusiva del SNP ha
La composicin en Ipidos de la mielina del
sido demostrada mediante sensibles tecni cas
SNP es similar a la del SNC, pero con diferencias
inmunohistolgicas en el SNC.
AORINA DEL PILAR LOPEZ Y HERNAN HURTADO GIRALDO
PAPELDELASCELULAS Al restablecer el contacto con los axones en
DE SCHWANN EN EL PROCESO regeneracin, las clulas de Schwann expresan
DE REGENERACION EN SNP Gal-C en su superficie y contraregulan los recep-
tores de NGF y molculas de adhesin (Cuadro
Mltiples evidencias experimentales, sugie-
3, 26,33). Si los axones alcanzan un dimetro
ren que las clulas de Schwann Son nicas en SU
suficiente paradisparar la mielinizacin, laclula
capacidad para estimular el Crecimiento y rege-
de Schwann es inducida a sintetizar MAG y
neracin del nervio perifrico. Dicha capacidad
aosteriormente POvarotenas bsicas de mielina
, .
persiste n v rro oonoe nan oemoslrado ser -n
14). Djrantc a suosec-enre m el nizacion \ a-
excelen~es~osrraroparae crec m enlooene-r' la~
r a s ~ l ~ asg a espar1 c pan en arem e n zacion
Las fases degenerativa y regenerativa en el
segmento distal del nervio perifrico lesionado
estn asociadas con extensos cambios en la
estructura y funcin de las clulas de Schwann.
Despus de la seccin del nervio, las clulas
gliales en el segmento distal sufren una fase
transitoria de intensa proliferacin, llevando a la
formacin de bandas de Bngner (8). Las clulas
mononucleares del torrente sanguneo entran en
la zona del trauma y ayudan a las clulas de
Schwann en el catabolismo de la mielina frag-
mentada y estimulan la proliferacin de estas,
probablemente por la secrecin de un factor de
crecimiento an no definido (52). La multiplica-
cin de lagladuranteladegeneracinwalleriana
comienza 1-5 das despus de la seccin del
nervio; el pico deactividad mitticaocurreaproxi-
rnadamente el da 30. y declina lentamente du-
rante las siguientes semanas. En el proceso de
degeneracin las clulas gliales del segmento
distal dejan de ensamblar lmina basal y de
expresar en su superficie celular el galactocere-
brsido (Cuadro 3. 29). Los niveles de m-RNA
para ciertas protenas caen dramticamente en
estas clulas y la pequena cantidad de POque es
sintetizada distalmente es degradada rpida-
mente en los lisosomas. La expresin de mol-
culas de adhesin de superficie se incrementa y
aumenta la expresin de receptores de baja
afinidad para el factor de crecimiento nervioso
(NGF. Cuadro 3). Los fibroblastos en el segmen-
to distal incrementan la produccin de colgeno
intersticial (42), los macrfagos que migran a la
lesin sintetizan y secretan interleucina 1 que
produce una rpida y transitoria elevacin de la
sntesis de NGF (52).
del que fuera previamente territorio de una sola,
acortando los internodos, loque asu vez produce
una marcada disminucin en la velocidad de
conduccin nerviosa. Durante la regeneracin
las clulas reexpresan N-CAM favoreciendo la
interaccion clula de Schwann-axn (43)
CELULAS DE SCHWANN IN VlTRO
UTILIDAD DE LOS CULTIVOS
Las tcnicas de cultivo de tejidos, ofrecen
posibilidades nicas para el estudio del desarro-
llo de las clulas de Schwann y para el anlisis
de las complejas interacciones celulares en el
SNP. Se han descrito diferentes mtodos de
cultivodepoblacionesdeclulasde laglaperifrica,
ya sea aisladas o asociadas con neuronas sen-
soriales. Como el ganglio sensorial madura in
vitro esto permite recapitular mucho de los estadios
del desarrollo como son el crecimiento de las
fibras nerviosas fuera del explante, la migracin
y la proliferacin de las clulas de Schwann a lo
largo de la fibra y los procesos de envolvimiento
y mielinizacin (44). Este sistema de cultivo hace
posible el anlisis de cada uno de los mltiples
factores que intervienen en cada fase. Adems
el cultivo permite trabajar en circunstancias que
hacen a las clulas acsequibles a laobservacin
directa y a la manipulacin.
La disponibilidad de los cultivos de clulas de
nervio perifrico en varias combinaciones ha
permitido por ejemplo el estudio de los origenes
de la matriz extracelular del nervio perifrico. Se
ha demostrado claramente que las clulas de
Schwann pueden sintetizar, secretar y ensam-
blarabundantescantidadesde matrizextracelular
(7). As mismo, el desarrollo de las tcnicas de
LA CELULA DE SCHWANN
cultivo ha permitido el estudio de la produccin
de factores de crecimiento, expresion de recep-
tores y molculas relacionadas con los procesos
de degeneracin y regeneracin posteriores a
una lesin de nervio perifrico. Otro aspecto
importante de los estudios in vitro con clulas de
Schwann es el haber permitido un anlisis de la
respuesta de dichas clulas a agentes infeccio-
sos como el Mycobacter~urn leprae a lo cual
haremos mencin ms adelante.
PATOLOGIAS QUE IMPLICAN
A LA CELULA DE SCHWANN
La vitalidad y funcionalidad de la clula de
Schwann como parte del nervio perifrico puede
verse afectada por mltiples factores de origen
diverso: infecciosos, inmunes, traumticos, txi-
cos y tumorales.
Dentro de los factores infecciosos se desta-
can el Mycobacteriurnlepraey el Corynebacterium
diphtheriae. Estos microorganismos causan en
la clula de Schwann alteraciones an no com-
pletamente aclaradas y que son todava objeto
de amplia investigacin.
-
El M. leprae es una micobacteria intracelular,
con una marcada afinidad por las clulas de
Schwann y del sistemaretculoendotelial(44)
Laentradadel M. IepraealacluladeSchwann
es un modelo nico de neurotropismo y es un
paso importante en la iniciacin de la cascada
de eventos que resultan finalmente en la
neuropatia leprosa (45). Estudios in vitro han
permitido comprobar que en cultivos de clu-
las de Schwann, neuronas y fibroblastos, el
M.lepraees fagocitado preferencialmente por
las primeras, encontrandose que detiene la
proliferacin de estas clulas y la subsecuen-
te mieiinizacin de los axones (45,46,47).
- La infeccin por el C. diphtheriae puede
causar una neuropata perifrica que se ca-
racteriza por la vacuolizacin y fragmentacin
de las vainas de mielina en el SNP. Los
hallazgos en modelos animales a los que se
les inyect la toxina producida por la bacteria,
sugieren que esta toxina parece actuar por
inhibicin de la sntesis de protenas de las
clulas de Schwann (26). La lesin caracteris-
tica es la desmieliniza-cin con preservacin
de la continuidad axonal, con lesiones
internodales en parche y una marcada prolife-
racin de clulas de Schwann (48).
Entre las alteraciones metablicas se desta-
ca la neuropatia diabtica, la ms comn de las
neuropatias perifricas, en la cual las clulas de
Schwann presentan una excesiva acumulacin
de cuerpos lipdicos en su citoplasma. Esta acu-
mulacin probablemente es reflejo de una altera-
cin en el metabolismo de los Ipidos y se produ-
ce una desmielinizacin segmentaria paranodal,
con formacin de bulbos de cebolla indicativos
de repetidadesmielinizacin (49,50). No sesabe
si la desmielinizacin es de tipo primario, o se-
cundaria a una alteracin axonal. Debido a la
presencia de aldoreductasa en las clulas de
Schwann, la hiperglicemia causa una excesiva
acumulacin desorbitol. Este pareceactuarcomo
una toxina tisular implicada en la patognesis de
la neuropatia (49,50,52)
La patologa tumoral que afecta las clulas de
Schwann es de caracter aeneralmente beniono
y se haclasificadoen cuatrogrupos: Schwannomas,
neurofibromas, fibromas plexiformes y fibromas
malignos. Cada uno de estos tipos puede verse
afectado en su conducta por la presencia de
neurofibromatosis. Los tumores de clulas de
Schwann son raros excepto en dos circunstan-
cias. La primera es la formacin no hereditaria,
espontnea, unilateral de un Schwannoma so-
bre el nervio vestibular que es llamado neuroma
acstico. La segunda consiste en un desorden
hereditario conocido como neurofibromatosis con
dos tipos bsicos: de Von Recklinghausen o
perifrica, caracterizada por mltiples neurofibro-
mas cutneos (tumores mixtos de clulas de
Schwann y fibroblastos) y la neurofibromatosis
acstica bilateral o neurof~bromatosis central dis-
tinta de la anterior pero tambin con carcter
autosmico recesivo (53). As mismo hay una
amplia lista de alteraciones inmunes metabli-
cas que modifican las clulas de Schwann Ile-
vando a procesos desmielinizantes que son ge-
neralmentesecundarios a lesion axonal queseran
motivo de otra revisin.
ADRINA DEL PILAR LOPEZ Y HERNAN HURTADO GIRALDO
SUMMARY production by Schwann cells. Ann. Rev. Neurosci.
1986; 9:305.
The Schwann cells, the glial cells of the
peripheral nervous system, suppori the neurona1
functions in several ways. Schwann cells are
implied infunctions like metabolicaxonal support,
facilitation of conduction and regeneration of
damaged axons.
Jessen KR, and Mirsky R. Schwann cell precursors
and their deveioprnent. Glia. 1991:4:185.
Komiyama A. and Susuki K. Age reiated difierences
in proliferative response of Schwann cells during
wallerian degeneration. Brain Res. 1992;576:267.
Bray GM, Rasminsky R, et al. lnteractions between
axons and their sheat cells. Ann. Rev. Neurosci.
1981;4:127.
In this paper we briefly review the origin of
these cells, their development, morphological
features, proliferation, mechanism of myelin for-
mation and the importante of the relationship
between axons and Schwann cells. At the end of
thisreview, we presentthemostcommondiseases
which affect the survival and normal function of
these cells.
REFERENCIAS
1. Duphin E, Baroffio A, Dulac C, et al. Schwann cell
diferentiation in conal cuitures of the neurai crest is
evidenced by the anii-Schwann cell rnyelin protein
monociona antibody. Proc. Natl. Acad Sci. USA.
1990:87:1119.2- Smith-Thomas LC and Fawcett JW.
Expression of Schwann c el markers by mamrnalian
neural crestcells i r vitro. Development.1989;105:251
3. BungeMB,WilliarnsAK, WoodPM.NeuronSchwann
cei interaction in basal lamina formation . Dev. Biol..
1982; 92:449.
4. Webster H. Developmentof Peripheral Nerve Fibers.
In Dick-Thomas eds. Peripheral Neurophaty. 1992;
Third Edition. Saunders Ed. Vol. 1 Pg.243.
5. Kelly BM,Guillespie CS, Sherman DLetal. Schwann
cells of the Myelin-forming phenotype express
NeurofilamentProteinNF-M.J.Cell Bioi. 1992; 118:397.
6. Salzer JL, and Bunge RP. Studies of Schwann cell
proliferation. l . An analysis i r Tissue culture of
proliferation during developrnent, Wallerian degeneration
and direct injury. J. Cell Biol. 1980:84:739.
7. Clark MB and Bunge ME. Cuitured Schwann cell
assernble normal-appearing basal lamina only when
they ensheate axons. Dev. Biol. 1989; 133:393.
8. Terzis JK and Smith KL. The Peripheral Nerve.
Structure Function and Reconstruction. Chapter 1
Raven Press. New York. 1990. Raven Press.
9. Jacobson M. NeuroglialOntogeny. In Developmental
Neurobiology. Chapter3.Third Edition. Penurn Press.
New York. 1991.
10. Bunge RP, Bunge M, and Elridge Ch. Linkage
betweeen axonal ensheiment and basal lamina
Salzer MJ, Pedraza L. Brown M, et al. Structure and
function ofthemyelingassociatedglycoproteins.Ann.
N. Y. Acad. Sci. 1990;650:302.
Ratner M, Bunge R. and Glaser L. Schwann cell
proliferationinvitro.Ann.N.Y.Acad.Sci.1986;486:170.
Davis J. and Stroobant P. Platelet-derived growth
factors and fibroblast growth factors are mitogensfor
rat Schwann cells. J. Ceil Biol. 1990;110:1353.
Lemke G, Kuhn R, Monuki ES, et al. Transcriptional
controls underlying Schwann cell diferentiation and
myeinitation. Ann. N. Y. Acad. Sci. 1990:605:248.
Muir D, Varon S, and Manthorpe M. Schwann cell
proliferation is under negative autocrine control. J.
Ceil Biol. 1990:111:2666.
Yoshi no JE. Mason PW. and DeVri es GH.
Uc c C- nci i a Ci oi qes nM/e 1- na..ccoprc 'cra:ois
? ' c . i .re'. S'i S . Ce 6 0 'S8' 102 655
Komivama A. and Susuki K. Aoe related chanaes i r
- ~ -
attachment and proliferation of mouse Schwann ceils
in vitro. J.Neurosc.Res. 1991;29:308-312.
Askanas V, Engel K, et al. Human Schwann cells in
tissuecuiture. Histochemicaland ultrastructuralstudies.
Arch. Neuro1.1980;37:329.
Dubois-DalcqM, Rentier B, BaronA.ef al. Sir .ci..rz
ar o i>e'.i 3' L' ld' C'i1.3 , 3 111 SFCCIILoI, sC1.'.311'
:e S i- . :,a 3. C? Res 168' ' ? 1 283 2: ?- Sr cce
ChF, Bunge ME, Bunge RP, et al. ~i fferenti ati onof
axonal-related Schwann cel s in vitro. J. Ceil Bioi.
1987;105:1023.
RossMH,and Romrell LJ. Eds. HistoIogyAtext and
atlas. Second edition, 1989: NervousTissue. Chapter
Goul d RM. Metabolic Organizatlon ofthe rnyeinating
schwann Cells. Ann. N.Y. Acad. Sci. 1990; 605: 44.
Morrel P, Quarles RH and Norton WT. Formation
structure and biochemistry of myelin. In Basic
Neurochemistry Text, Fourth edtion, Raven Press.
New York. 1989.
27. Bunge ME, Williams A, Wood PM, etal . comparison
of nerve cells and nerve cell ~ l u s Schwann ceils
cultures with particuiaremphasis'on basa laminaand
collayen formation. J. Cell Biol. 1980; 84: 184.
28. Bunge ME, Deanet AC, et al. Schwann cell function
depends upon axonal sgnai s and basal lamina
components. Ann. N. Y. Acad. Sci. 1990: 580: 281
29. Carey DJ, Elridge ChF, Cornbrooks CJ, et al.
Byosynthesis of type IV colagen by Rat Schwann
ceils. J. CeI Bi ol 1983;87:473~479.
30. Lemke G. Unwrappiny the yenes of myelin. Neuron.
1988:1:535~543.
3' Mo6uno K.Kamholz J. Behrman T. el al. I \ e . . r ~ i o
- 2' . . r. o. i 3i Sc, . . a~r :e s ' r r ar, o: o :p.oie n
U-PP . I\a .rcsc H?s 1569 23 39~i - i C5
3: U' .'. e q i SM .g 1. 1 I A ,r- Era,, ST P ? _ccn
' )'.. . II onne.? al"eii--ccnr Sr i i ~ U' ? =:id C? 3;'
and slow axonai transp& by myelinatiny Schwann
cells. Cell. 1992;68:451-463.
33. Eun-hye J. and Ki mon A. Clusterinu of voltaae~
41.
Bi xby JL, Lilien J, and Reichardt LF. ldentification
of the majar proteins that promote neurona1 process
outyrowth on Schwann cells in vitro. J.Celi Biol.
198;107:353.
42. TaniuchiM,ClarkHB,SchweitzerJB,etal.Expression
of neme arowth factor receotors bv Schwann ceils of
u
axotomized peripherai nerves. Ultrastructurai location.
supression by axonai contact and bindiny properties.
J.Neurosci. 1988:8.2:664.
43. Rieqer F, Nicolete M. Pincon-Ravmond M. et al.
~i st i bi t i on and role in regenerationof N-CAM in the
basal laminae of muscle and Schwann cells. JCel l
Biol. 1988;107:707.
44. MehtaR. BirdiTJand AnthiaNH. Effectofthe M.ieprae
infected Schwann ceils and their supernatant on
lymphocyte neuroglia interaction. JNeuroimmunol.
1989,22:149-155.
45. Job CK, and Veryhese P. Schwann cells chanyes in
lepromatousieprosy. Indian J.Med.Res.l975;63:879.
46- Jacobs JM. Shetty VP. and Anthia NH. Myelin
chanues in IeDrous neurooathv. Act. Neuroohat.
dependent sodium chanels on axons'depends-on 1987-/4:75
Schwann cell contact. Nature. 1992;356:333-335.
47. MahadevanPR,andAntiaNH.Biochemicalalterations
34. Rosenbluth J. Roie of glial ceils in the differentiation
in cells foliowing phagocytosis of M. leprae -The
an f unct on of myel i nated axons. 1nt.J. consecuense- A basic concept.lnt.J.of Leprosy.
Dev.Neurosc.l988;6:1:3. 1980.48;2:167.
3 5 Filbin MT, Walsh Fs, Trapp BD, et 48. McDonaldW.I.andKocenRS.Diphterieticneurophaty.
protein as an homophlic adhesion molecuie. Nature.
i n Dick-Thomas eds. Peripheral Neurophaty. Third
1990;344:871
edition. 1992 vol. 2 Chapter 77.Pag.1412.
36. Wood WP,Moya F, Eldridge Ch. et al. Studies ofthe
i ni t at i on of mMyel i nat on by Schwann cells.
Ann.N.Y.Acad.Scien. 1990;605:14.
37. Mezel C. Myelination in the peripheral nerve duriny
deveopmentln Dick-Thomas Ed. Peripheral Neurophaty.
Third Edition.1992. Vol1 Chapter 16. Pg. 267.
38. Owens GC, Boyd CD, Bunge RP, et al. Expression
ofrecombinantmveiin-associatedalicoorotein in Primar"
" , ,
Schwann celis promotes the nitial investmentofaxons
by myelinatiny Schwann cells. JCel i Biol. 1990; l l l
39. Mi rskv R. and Jessen K. The Biooov of Non- rnvelin
-,
f o r mi y ~ c h w a n n celis. ~ n n . ~ . ~ . ~ c a d . ~ c i e n
1986;486:132.
40. Selheimer B. and Schacher H. Reaulation of neuial
49. Mogensen CE, and Christensen CK. Predicting
diabetic neuropathy in insulin dependent patients.
N.Enyl.J.Med. 1984;311:89.
50. Thomas PK, and Thoml i nson DR. Diabetic and
Hipoylicernic Neuropathy. In Dick-Thomas eds.
Peripherai Neuropathy. Third Edition. 1992.Vo1.2.
Chapter 64. Py.1219.
51. Foster DW. Diabetes Mellitus. in Harrison'sPrinciples
of Interna1 Medicine. international Edition. Twelfth
Edition.1991. Vo l 2 Chapter 339. Pg.1739.
52. Caicutt NA, Muir 0, Powell HC,etal. Reduced ciliary
neurotrophcfactor iikeactivity in nervesfrom diabetic
or yalaciose fed rats. Brain Res. 1992;579:320.
u ~ ~
ceil adhesion moleculeexpression on cultured rnouse 53. Brockes JP, Breakefield XO and Martuza RL. Glial
Schwann cel s bvnerve qrowthfactor. EMBO Journal. ~r owt h factor-like activitv in Schwann cells tumors.
198766:1611 nn. Neur o 1986;20:31>
Vous aimerez peut-être aussi
- ERITROSEDIMENTACIONDocument6 pagesERITROSEDIMENTACIONJhoana Veronica Mariscal RochaPas encore d'évaluation
- HISTORIADocument7 pagesHISTORIAcibert4Pas encore d'évaluation
- Razonamiento Verbal 2º Año - I BimestreDocument7 pagesRazonamiento Verbal 2º Año - I Bimestrecibert4Pas encore d'évaluation
- Articulo Gen AhorradorDocument3 pagesArticulo Gen Ahorradorcibert4Pas encore d'évaluation
- Disposiciones Complementarias Proceso Admision Residentado Odontologico 2017Document21 pagesDisposiciones Complementarias Proceso Admision Residentado Odontologico 2017cibert4Pas encore d'évaluation
- Karl Landsteiner y Los Grupos SanguineosDocument3 pagesKarl Landsteiner y Los Grupos SanguineosCristian Martinez Carbajal0% (1)
- Colección de Archivos de Medicina: Año 2005Document221 pagesColección de Archivos de Medicina: Año 2005International Medical PublisherPas encore d'évaluation
- Proceso de Contraccion MuscularDocument11 pagesProceso de Contraccion MuscularramPas encore d'évaluation
- Laboratorio 9Document11 pagesLaboratorio 9Brenda Aguilar SaraviaPas encore d'évaluation
- Determinación de Grupo Sanguíneo Sistema ABO y RHDocument6 pagesDeterminación de Grupo Sanguíneo Sistema ABO y RHHector RojasPas encore d'évaluation
- Desarrollo Del Hueso y CartilagosDocument3 pagesDesarrollo Del Hueso y CartilagosStefaniaPas encore d'évaluation
- Fraccionamiento Automatizado E Vargas R Rodas Paraguay Sep 2020Document14 pagesFraccionamiento Automatizado E Vargas R Rodas Paraguay Sep 2020Daniel Apaza SolanoPas encore d'évaluation
- Trabajo de Estudio de La Serie RojaDocument5 pagesTrabajo de Estudio de La Serie Rojaesther barbaran rojasPas encore d'évaluation
- Histología de Los Tejidos Del CuerpoDocument13 pagesHistología de Los Tejidos Del Cuerpomichelle bernalPas encore d'évaluation
- Referencia CardioDocument17 pagesReferencia CardioAlex Menoscal BermeoPas encore d'évaluation
- Histología Del OvarioDocument6 pagesHistología Del OvarioVictor Pier Chuquipoma RojasPas encore d'évaluation
- Las Respuestas Fisiológicas en El Paciente Quirúrgico IDocument45 pagesLas Respuestas Fisiológicas en El Paciente Quirúrgico IClaudia AlessandraPas encore d'évaluation
- Anemia Severa Normocitica NormocromicaDocument4 pagesAnemia Severa Normocitica NormocromicaJose David Lopez Velasquez100% (1)
- Modelo de HillDocument17 pagesModelo de HillSantiagoPas encore d'évaluation
- Nivel Tisular Mapa ConceptualDocument1 pageNivel Tisular Mapa ConceptualFREDDY ELIAS PERILLA PORTILLAPas encore d'évaluation
- CoagulacionDocument10 pagesCoagulacionHernan GonzalezPas encore d'évaluation
- Muestra SanguineaDocument5 pagesMuestra SanguineaTracy Lopez BullonPas encore d'évaluation
- 1634 Presentación Electrónica Educativa 1646 1-10-20190313Document42 pages1634 Presentación Electrónica Educativa 1646 1-10-20190313SócratesNautoPas encore d'évaluation
- Evaluación de Los Trastornos Hemorrágicos en NiñosDocument40 pagesEvaluación de Los Trastornos Hemorrágicos en NiñosMarcela Elizabeth CevallosPas encore d'évaluation
- Pasitos SemanalesDocument9 pagesPasitos Semanalesvaleria torrealvaPas encore d'évaluation
- CoagulogramaDocument25 pagesCoagulogramaYailin Sánchez MesaPas encore d'évaluation
- Tejido CartilaginosoDocument5 pagesTejido CartilaginosoOscar PerezPas encore d'évaluation
- Contracción MuscularDocument6 pagesContracción MuscularStephanie GuerreroPas encore d'évaluation
- NERVIOSODocument7 pagesNERVIOSOEvelin SánchezPas encore d'évaluation
- Reporte Práctica 2 Inmuno SISTEMA ABO Y RHDocument6 pagesReporte Práctica 2 Inmuno SISTEMA ABO Y RHJosé Francisco Valenzuela SánchezPas encore d'évaluation
- Preprácticos Histología Cat2Document53 pagesPreprácticos Histología Cat2hnbaePas encore d'évaluation
- Tejidos Biológicos Estructura y Función FinalDocument67 pagesTejidos Biológicos Estructura y Función Finalmonko.y2kPas encore d'évaluation
- Practica No. 6 Recuento de Plaquetas Con HemocitometroDocument4 pagesPractica No. 6 Recuento de Plaquetas Con HemocitometroSandra RdzPas encore d'évaluation
- ANEMIA Medicina InternaDocument8 pagesANEMIA Medicina InternalissethPas encore d'évaluation