Académique Documents
Professionnel Documents
Culture Documents
Tonus Motricite
Transféré par
charbelhannaCopyright
Formats disponibles
Partager ce document
Partager ou intégrer le document
Avez-vous trouvé ce document utile ?
Ce contenu est-il inapproprié ?
Signaler ce documentDroits d'auteur :
Formats disponibles
Tonus Motricite
Transféré par
charbelhannaDroits d'auteur :
Formats disponibles
1
re
partie : Neuroanatomie fonctionnelle
Chapitre 9 TONUS ET MOTRICITE
La physiologie du mouvement comprend la motricit, les rflexes et le tonus musculaire. Les
mcanismes qui contrlent le mouvement sont complexes et imparfaitement connus. Le motoneurone de la
corne antrieure de la moelle et les fibres musculaires stries qui en dpendent reprsentent la voie finale
commune vers laquelle convergent les diffrentes influences qui vont venir modifier, moduler leur
fonctionnement. Il est habituel dans un but de simplification, d'envisager des niveaux d'intgration successifs.
I - Niveau mdullaire
Des activits motrices fondamentales sont dj inscrites dans la moelle. Elles reprsentent des mcanismes
rflexes : rflexes proprioceptifs en extension, rflexes nociceptifs en flexion. Pour ce qui concerne les rflexes, il faut
mentionner aussi les rflexes viscraux.
II - Niveau du tronc crbral
Ces activits sont asservies par le dispositif du tronc crbral qui mettent profit les affrences vestibulaires et
cutanes pour ordonner le redressement et les adaptations posturales.
L'adoption de la station rige ncessite un renforcement de l'activit des muscles antigravifiques
(c'est--dire des muscles extenseur physiologiques). Ce tonus antigravifique repose sur le rflexe
myotatique. Le dispositif essentiel qui assure de faon tonique l'activation du rflexe myotatique de muscles
antigravifiques et la substance rticule activatrice descendante, dont l'action est essentiellement assure par le
faisceau rticulo-spinal mdian. Ce mcanisme n'est pas le seul qui intervienne au niveau du tronc crbral et les
noyaux vestibulaires par l'intermdiaire de la voie vestibulo-spinal exercent une action facilitatrice sur le tonus des
muscles extenseurs.
Le dplacement actif ou passif d'une partie du corps par rapport l'axe de la vertical entrane une srie
d'adaptations automatiques du tonus musculaire en vue du maintien de l'quilibre. Ces adaptations
posturales des rflexes antigravifiques ncessitent une intgration des stimulations affrentes
proprioceptives, vestibulaires, cutanes, visuelles. Celle-ci se produit pour une part au niveau du tronc
crbral suprieur, mais elle n'est assure de faon dfinitivement efficace qu'au niveau thalamo-palidal.
III - Niveau crbral
L'tage le plus volu est reprsent par le cortex crbral et le no-striatum (noyau caud, putamen).
La fonction concerne ici et le contrle du mouvement propositionnel (tlognique), mouvement dirig vers le monde
extrieur qui peut revtir l'aspect d'une rponse automatique ou celui d'un mouvement volontaire .
Les noyaux gris centraux assurent la formulation globale de ce mouvement en intgrant et modulant les
acquisitions des niveaux sous-jacents. La participation autonome des noyaux gris centraux au mouvement
propositionnel est probable en raison du dveloppement considrable des relations entre striatum et cortex
crbral. Des ractions motrices fondamentales telles que la prhension, l'vitement, la rotation de la tte et des
yeux sont intgres ce niveau.
Le cortex crbral exerce une action trs gnrale sur la motilit. Les expriences de stimulation et de
destruction localise montre que cette proprit loin d'tre limite au cortex moteur pr-rolandique est tendue
de trs nombreuses rgions du cortex. Nanmoins une distinction s'impose entre la zone motrice principale que ces
effrences cortico-spinales (faisceau pyramidal) mettent directement en rapport avec les dispositifs segmentaires
de la moelle et les autres rgions qui n'exercent leurs effets moteurs qu'indirectement par l'intermdiaire des
formations grises sous corticaleS (cortex extrapyramidal) et de la substance rticule. La motilit qui persiste aprs
section du faisceau pyramidal est relativement riche mais elle est remarquable par son caractre global et
syncintique. Cette donne ainsi que l'troite relation fonctionnelle que prsente le cortex moteur principal avec
l'air parital o se terminent les voies de la sensibilit discriminative et avec le noyau ventro-latral du thalamus
laissent prsager du rle essentiel de ce cortex moteur principal dans les adaptations spatio-temporelles du
mouvement en fonction des qualits de la stimulation.
IV - Niveau crbelleux
Le cervelet plac en drivation sur le dispositif sensori-moteur qui rgle le mouvement n'est indispensable aucune
des formes de la motilit. Nanmoins la dfaillance du contrle crbelleux se marque dans chacune de ces activits par la
perte de leur coordination harmonieuse.
Une telle division en secteurs tags constitue cependant une simplification excessive qui ne doit pas masquer
l'extrme complexit des mcanismes qui contrlent le mouvement (fig. 1).
120
L'unit motrice - syndromes neurogne priphrique et syndrome myogne
L'ensemble form par un motoneurone alpha et les cellules musculaires stries qui en dpendent constitue une
unit motrice, voie finale commune de la motilit.
I -structures et fonctionnement de l'unit motrice (fig. 2)
le motoneurone : son corps cellulaire est situ dans la corne antrieure de la moelle. Son axone, fibre mylinise
de gros diamtre et de conduction rapide, se ramifie la surface de la cellule musculaire.
La cellule musculaire strie se caractrise essentiellement par l'existence d'un double systme de rticulum
endoplasmique lisse et par la prsence d'un matriel contractil : les myofibrilles. Chacune, faite de la succession
rgulire de cylindres identiques appels sarcomres, comporte deux types myofilaments (actine et myosine) dont
la rpartition au sein des sarcomres dtermine des rgions de structures diffrentes responsables de la striation
transversale. Suivant certaines caractristiques morpho-fonctionnelles, on distingue les fibres de types I (muscles
rouges contraction lente), de types II (muscles blancs contraction rapide, forte activit ATPasique) et de type
intermdiaire.
Fig. 1 - Diagrammes des systmes anatomo-fonctionnels contrlant la motricit.
Chaque niveau de contrle et les structures qui en dpendent sont reprsentes par une couleur diffrente. Notez la
terminaison directe des influences vestibulaires sur le motoneurone
La plaque motrice (ou jonction neuro-musculaire) permet la transmission de l'influx nerveux la cellule musculaire.
Il s'agit d'une structure composite appartenant en partie au nerf moteur, en partie la cellule musculaire. ce
niveau l'arborisation terminale de l'axone repose dans une gouttire directement sur la membrane de la cellule
musculaire donc il n'est spar que par la fente synaptique. Cette membrane prsente ce niveau de multiples
invaginations parallles qui constituent l'appareil sous-neural, trs riche en cholinestrase.
Au sein de l'unit motrice, la dpendance de la cellule musculaire vis--vis du motoneurone alpha est dmontre
par la section nerveuse qui entrane son atrophie (atrophie de dnervation). Celle-ci est prcde d'une
dgnrescence des plaques motrices. Par ailleurs, une unit motrice est forme de cellules musculaires du mme
type. Il apparat ainsi que la cellule nerveuse exerce une influence permanente sur les cellules musculaires qu'elle
commande.
Fig. 2- Schma de la constitution d'unit motrice
121
Fonctionnement de l'unit motrice
Toute dcharge d'un neurone moteur active l'ensemble des fibres musculaires qui constituent l'unit motrice.
Transmission de l'excitation de la cellule musculaire strie
Elle se fait suivant le mode gnral de la transmission synaptique : la dpolarisation de la membrane prsynaptique
qui traduit la stimulation axonale dclenche la libration d'actylcholine dans la fente synaptique partir des
vsicules synaptiques. L'actylcholine modifie la permabilit aux ions Na
+
et K
+
de la membrane plasmique de la
cellule musculaire formant l'appareil sous neural et en provoque la dpolarisation (potentiel de plaque motrice) qui,
si elle est d'amplitude suffisante, est l'origine d'un potentiel propag. La prsence de cholinestrase en excs
assure la rversibilit instantane de l'action du transmetteur, de telle sorte que la plaque motrice au repos
l'arrive est disponible nouveau.
Par ailleurs, l'enregistrement prolong de l'activit lectrique d'une plaque motrice en l'absence de toute
stimulation du nerf, met en vidence des potentiels de plaque miniature (mepps) qui correspondent la libration
irrgulire spontane du contenu d'une vsicule synaptique (quanta de neurotransmetteur).
La contraction musculaire.
La contraction des myofibrilles rpond la modification des liaisons assurant filaments d'actine et de myosine. Il en
rsulte un glissement des uns et des autres avec raccourcissement global des sarcomres. Cette modification
structurale est associe une dephosphorylation de l'ATP musculaire et est troitement dpendante des
modifications de la concentration en ions Ca
2+
du rticulum sarcoplasmique.
Smiologie de l'unit motrice
Les perturbations de l'activit de l'unit motrice peuvent intresser :
-- son versant nerveux : lsion du neurone moteur (syndromes neurognes priphriques) ;
-- son versant musculaire : altration primitive de la fibre musculaire (syndrome myogne) ;
-- la jonction neuro-musculaire perturbations de l'unit motrice rsultant d'une dgradation de l'efficacit synaptique la
jonction neuro-musculaire. Dans des blocs neuro-musculaires existe un contraste entre le dsordre majeur du
fonctionnement et l'intgrit anatomique relative de l'unit motrice.
Les rflexes
Un rflexe est une rponse motrice involontaire une stimulation sensitive ou sensorielle. L'activit rflexe intgre
un niveau segmentaire est soumise au contrle des structures sus-jacentes. L'examen des rflexes est donc un test qui
value un moment prcis la ractivit du systme nerveux dans son ensemble.
Organisation physiologique
Le support anatomo-fonctionncl de la liaison qui unit un stimulus spcifique une rponse dtermine est un arc
rflexe.
1. Les rflexes d'origine proprioceptive
Le rflexe monosynaptique ou myotatique correspond la varit phasique du rflexe osto-
tendineux (fig. 3).
Fig. 3 Exemple de rflexe monosynaptique: le rlexe myotatique
Le rcepteur form par la terminaison primaire du fuseau neuro-musculaire est sensible l'tirement. Par
l'intermdiaire des fibres la conduction rapide du fuseau, qui cheminent dans le nerf priphrique, la vole affrente est
transmise la moelle o par une seule synapse elle gagne les motoneurones "commandant les fibres musculaires d'o vient
l'excitation".
Plusieurs systmes modulent l'efficacit du rflexe myotatique par contrle du niveau d'excitabilit des
motoneurones.
Les motoneurones . situs au voisinage des prcdents dans la corne antrieure de la moelle, innervent la partie
contractile du fuseau neuro-musculaire et dterminent sa sensibilit l'tirement. Ces motoneurones sont contrls de
faon indpendante par les structures supra-segmentaires. Dans le circuit rcurrent inhibiteur de Renshaw, l'axone du
122
motoneurone donne une collatrale rcurrente qui s'articule avec un interneurone. Celui-ci inhibe le motoneurone ainsi
que les neurones moteurs synergiques voisins vitant ainsi une synchronisation excessive de l'activit des motoneurones.
D'autres rflexes sont sous la dpendance des affrences proprioceptives.
Innervation rciproque : les mmes affrences qui dterminent le rflexe monosynaplique ont une action inhibitrice par
l'intermdiaire d'un interneurone sur les muscles antagonistes du muscle tir. Ainsi toute modification dans la frquence
des influx transmis par les fibres la d'un muscle donn retentit ncessairement sur les motoneurones de ce muscle, des
muscles synergiques et des muscles antagonistes. A l'ensemble des muscles d'une articulation donne qui sont ainsi
mutuellement dpendants on a donn le nom d'unit mvotatique.
Rflexe mvotatique inverse : l'activation des organes tendineux de Golgi transmise par les fibres Ib inhibe, par
l'intermdiaire d'un interneurone, les motoneurones des muscles synergiques et facilite au contraire les motoneurones des
muscles antagonistes. Cette raction correspondrait un rflexe de protection mettant le muscle l'abri de la rupture en
cas de tension excessive.
Rflexes de posture : il s'agit de la contraction qui survient dans un muscle aprs l'arrt du mouvement passif de
raccourcissement de ce dernier. Le rle et les voies de ces activits rflexes sont mal connus.
2. Les rflexes d'origine cutane
L'arc rflexe est toujours polysynaptique : les affrences sont vhicules par des fibres d'origine cutane (diamtre
infrieur celui des fibres Ia) et plusieurs interneurones s'interposent entre elles et les motoneurones , voie effrente
oblige.
Les rflexes polysynaptiques dclenchs les par des stimuli nociceptifs sont des rflexes en flexion finalit
protectrice. La raction de retrait qui en rsulte est influence par le sige de la stimulation (fig. 4).
Les rflexes polysynaptiques dclenchs par des stimuli non nociceptifs ont une organisation est une signification moins bien
connue.
La smiologie des rflexes comprend leur tude, et l'examen de leurs anomalies.
Fig. 4 Exemple de rflexe polysynaptique: le rflexe de retrait d'un membre enrponse un stimulus
nociceptif
La voie motrice principale : le syndrome pyramidal.
Au sens strict le faisceau pyramidal dsigne l'ensemble des fibres qui constituent la pyramide bulbaire mais sous le
terme de syndrome pyramidal on groupe un ensemble de troubles moteurs et rflexes qui traduit la lsion des fibres
empruntant le trajet de ce faisceau sur le sa totalit ou en partie. L'htrognit anatomo-fonctionnelle du faisceau
pyramidal tant clairement dmontre, il est sr que le syndrome pyramidal ne correspond pas une lsion touchant
uniquement les fibres pyramidales.
1. La voie pyramidale
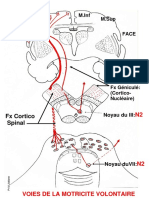
Organisation anatomique (fig. 5)
La totalit des fibres composant le faisceau pyramidal provient de neurones situs dans le cortex crbral. Toutefois
la contribution des aires motrices (frontale ascendante, ou aire 4 et aire prmotrice, ou aire 6) n'est que d'environ 40 %. Un
autre contingent vient des aires paritales mais 30 40 % des fibres prennent naissance ailleurs que dans les aires
centrales.
Au niveau de la circonvolution frontale ascendante existe une disposition somatotopique que reproduit l'homonculus
de Penfield : la face, la main et le pouce occupent une place considrable.
Le cortex moteur contrle la musculature contralatrale mais pour l'extrmit cphalique et la musculature axiale
du cou et du tronc, l'existence d'une reprsentation bilatrale est certaine.
partir de leur origine corticale, les fibres pyramidales descendent dans le centre ovale puis la capsule interne.
ce niveau le faisceau cortico-bulbaire, destin aux noyaux somato-moteurs des nerfs crniens est en avant du faisceau
cortico-spinal.
Dans le tronc crbral, le faisceau pyramidal occupe les 4/5 internes du pied du pdoncule puis est dissoci par les
fibres ponto-crbelleuses dans le pilier de la protubrance. Regroupdans la pyramide bulbaire, il subit dans la partie basse
du bulbe un entrecroisement partiel qui intresse 80 % des fibres.
Dans la moelle le faisceau pyramidal crois descend dans la partie postrieure du cordon latral et le faisceau
pyramidal direct, plus grle, dans le cordon antrieur.
Le faisceau cortico-bulbaire (gnicul) s'puise diffrents niveaux du tronc crbral en donnant des fibres croises
et directes aux noyaux moteurs des nerfs crniens. De nombreuse collatrale sont destines la substance rticule.
La plupart des connexions avec les neurones moteurs priphriques ne se font pas directement, mais par
l'intermdiaire d'un systme de neurones intercalaires. Les connexions cortico-motoneuronales directes empruntent
probablement au sein du faisceau pyramidal le contingent de grosses fibres mylinises conduction rapide.
123
Physiologie lmentaire
La voie pyramidale apparat comme un chanon indispensable du systme contrlant certaines formes privilgies
de mouvements volontaires notamment les mouvements finement ajusts de la main et des doigts. Les influx cortico-
spinaux moduls au niveau cortical par de multiples affrences permettent la fragmentation des synergies primitives en
mouvements isols. Ils assurent la slectivit du mouvement en inhibant les synergies inscrites l'chelon mdullaire
segmentaire entre les diffrents muscles.
Les modifications rsultant d'une interruption du faisceau pyramidal dpendent chez l'animal du niveau de la lsion:
Une destruction corticale (aire 4) comme l'interruption du faisceau pyramidal au niveau de la pyramide
bulbaire donne lieu une hmiplgie flasque. L'hypertonie d'un type particulier qui est le symptme
majeur du syndrome pyramidal clinique ne fait donc pas partie de cette varit particulirement pure du
syndrome pyramidal exprimental. Il est possible qu'elle rsulte de l'interruption de fibres cortico-
rticulaires (soit distinctes, soit collatrales des axones pyramidaux) doues d'une action inhibitrice sur la
formation rticule activatrice descendante. Cette dernire, prive de ses influences inhibitrices, exercerait
par des voies descendantes extrapyramidales, une action excitatrice sur les mcanismes rflexe
segmentaires proprioceptifs.
Au cours des atteintes mdullaires, l'hypertonique spastique est souvent trs accuse. Ceci tient
probablement au fait que les influences facilitatrices s'exerant sur les motoneurones passe par le
faisceau rticulo-spinal mdian qui est le faisceau descendant le plus diffus et donc le plus difficile
interrompre.
Fig. 5 Anatomie du faisceau pyramidal
2. Smiologie analytique du syndrome pyramidal
L'hypertonique pyramidale
Marque au minimum par une exagration des rflexes ostotendineux, la spasticit est le symptme essentiel du
syndrome pyramidal.
L'hypertonie spastique se manifeste lors de la mobilisation passive qui tire un muscle, sous la forme d'une
contraction rflexe qui s'oppose l'tirement qui lui a donn naissance.
La spasticit prdomine toujours et souvent n'existe que sur l'un des lments des diffrents couples musculaires
agoniste-antagoniste qui agissent sur une articulation donne.
L'hypertonie spastique et l'expression clinique de l'exagration du rflexe myotatique d'tirement.
Les modifications des rflexes cutans
L'inversion du rflexe cutan plantaire constitue le signe de Babinski.
On observe aussi l'abolition de certains rflexes polysynaptiques non nociceptifs.
Certains rflexes d'origine cutane sont l'inverse facilits par la lsion de la voie pyramidale.
Les difficults de commande volontaire
En cas de lsion aigu et rcente, il existe souvent au dbut une impossibilit totale de toute commande motrice
volontaire dans le territoire correspondant.
La rpartition du dficit moteur est globalement inverse de celle de l'hypertonie.
Le dficit prdomine aux extrmits distales et sur la motilit labore de la face.
124
Les syncinsies
Les syncinsies de coordination consistent dans le fait que la contraction volontaire de certains groupes musculaires
entrane la contraction involontaire syncintique de groupes musculaires fonctionnellement synergiques.
Les syncinsies globales sont dclenches par un effort effectu contre rsistance par un membre sain et tendent
reproduire en l'exagrant la topographie de l'hypertonie.
3. Syndrome pyramidal et niveau lsionnel
Au cours des atteintes encphaliques, un seul faisceau pyramidal est touch. Les troubles observs n'intressent
que un hmicorps : c'est l'hmiplgie qui est croise par rapport la lsion.
Dans les atteintes mdullaires, les voies motrices correspondant aux deux cts sont en gnral touches du fait de
leur proximit : c'est la paraplgie ou ttraplgie.
Le syndrome pseudo bulbaire est la consquence de lsions bilatrales du faisceau gnicul. Il se caractrise par un
tat parto-spasmodique touchant la face, la langue, le pharynx, le larynx et les masticateurs.
Le systme extrapyramidal
La smiologie extrapyramidale groupe l'ensemble des dsordres de la motilit qui rsulte de la perturbation du
fonctionnement des dispositifs extrapyramidaux, et en particulier des noyaux gris centraux.
1 - Organisation anatomo-fonctionnelle du systme extrapyramidalL
Les dispositifs extrapyramidaux (fig. 6 )
Fig. 6 Le systme extra-pyramidal
La complexit anatomique du systme extrapyramidal s'oppose l'organisation relativement simple du systme
pyramidal. De nombreuses structures en font partie : au niveau du cortex de crbral, la rgion prmotrice situe en avant
de l'aire 4; parmi les structures profondes, le striatum, le pallidum et certains noyaux du thalamus qui constituent le
dispositif central du systme ; au niveau de la partie haute du tronc crbral, le noyau sous-thalamique, le locus niger et le
noyau rouge qui compltent les formations thalamo-stries et ; enfin la substance rticule.
Les relations qui existent entre ces diffrentes structures sont complexes et encore mal connues. Multiples, souvent
bilatrales, elles forment des circuits qui se contrlent mutuellement tout moment.
Une schmatisation l'extrme permet d'envisager un dispositif central, le systme strio-palidal, avec son double
contrle interne assur par le locus niger et le noyau sous-thalamique. Ce systme reoit tout moment des informations
venant du cortex prmoteur et de la substance rticule. En change il envoie des informations vers la substance rticule
et par le thalamus vers le cortex prmoteur (boucle cortico-thalamo-corticale). Celui-ci outre son influence sur le systme
strio-palidal projette sur les niveaux sous-jacents par l'intermdiaire du faisceau pyramidal et de leur substance rticule.
La substance rticule reprsente ainsi l'aboutissement de nombreuses voies venant de diffrents niveaux du
systme extrapyramidal. D'elle, partent des faisceaux projection bilatrale qui vont moduler l'activit des motoneurones
et .
Spcificit fonctionnelle du systme strio-pallidal
Corps stris et thalamus forment le centre des dispositifs extrapyramidaux. Une spcificit certaine existe dans le
fonctionnement des diffrentes structures. Ainsi, le striatum, intgrateur des affrences thalamiques et corticales, exerce
une action inhibitrice globale sur les mcanismes moteurs et donc probablement sur le pallidum. Celui-ci, effecteur
125
obligatoire du systme, a une action excitatrice importante sur ces mmes mcanismes. Le locus niger et le noyau sous
thalamique sont un frein inhibiteur majeur du fonctionnement respectif des striatum et du pallidum. Quant au thalamus, il
est la fois une des sources principales des affrences du systme (noyau centre mdian) et les premiers relais des
projections strio-palidales vers le cortex. C'est au niveau de son noyau ventro latral que se fait la convergence des
informations extrapyramidales et crbelleuses. L'importance de cette convergence a t rendue manifeste par le succs
dans certains troubles de la motilit de la chirurgie strotaxique dont ce noyau est la cible privilgie.
Les tudes biochimiques ont permis de prciser certains aspects de cette spcificit. Le striatum est riche en
actylcholine et en dopamine, substances qui exercent sur son fonctionnement une activit respectivement excitatrice et
inhibitrice. Il a t dmontr que le contrle dopaminergique du striatum s'exerce par des neurones nigro-stris dont le
corps cellulaire et situ dans le locus niger, structures galement trs riches en dopamine. Il est probable que ces neurones
s'articulent dans le striatum avec des neurones cholinergiques, et que ces derniers agissent sur les neurones effecteurs du
striatum dont le neuromdiateur pourrait tre le GABA (fig. ).
La connaissance des neuromdiateurs et des chanes neuronales qui les utilisent au niveau des noyaux gris restent encore
fragmentaires. Elle est d'ores et dj riche de consquence dans le traitement d'affections extrapyramidale.
Fonction des noyaux gris centraux
L'organisation anatomique des dispositifs extrapyramidaux explique la formulation trs globale de leur action sur
l'ensemble de l'conomie. Ils assurent dans le contrle du mouvement une triple action :
couronnant les mcanismes du tronc crbral et intgrant les informations d'origine vestibulaire et
visuelle, ils achvent l'acquisition de la station rige ;
modulant ces activits toniques fondamentales et leur superposant des activations phasiques, ils rendent
possibles les adaptations posturales ;
sous le contrle du cortex extrapyramidal, ils organisent les rponse motrices globales aux stimulations
extrieures et sous-tendent l'activation de la voie cortico-spinale laquelle sont dvolues les adaptations
fines du geste.
2 -Smiologie extrapyramidale
Le dysfonctionnement du systme extrapyramidal n'entrane pas de paralysie mais des perturbations toniques et
des modifications de l'activit motrice qui peuvent se faire dans le sens d'une diminution (akinsie) ou d'une augmentation
(hyperkinsie).
En dehors de quelques exceptions, la valeur localisatrice d'un signal appartenant cette smiologie est faible.
Rsultat en effet de la perturbation d'un systme de contrle complexe, son origine est chercher dans les conditions
d'quilibre qui se sont reforms aprs lesion d'un ou de plusieurs lments de ces circuits. Des lsions de topographie
diffrente peuvent entraner une perturbation motrice du mme ordre, probablement parce qu'elles modifient dans le mme
sens l'quilibre des circuits engags dans le mouvement.
Le syndrome parkinsonien
L'akinsie se dfinit par la raret du geste et par les difficults prouves par un sujet raliser la moindre
performance motrice. Trouble de l'initiation et de l'excution du mouvement, elle se manifeste la face (aspect fig du
visage) et aux membres (disparition des mouvements spontans). La perte des mouvements automatiques et associs
(balancement des bras la marche par exemple) est la base des perturbations observes dans le mouvement volontaire
et qui se traduisent par un retard l'initiation motrice, un droulement ralenti et hsitant du geste, une impossibilit
d'excuter des activits conjugues (marionnettes).
Composantes ngatives du syndrome parkinsonien, l'akinsie traduit l'activit inhibitrice excessive du striatum sur
le pallidum activit qui n'est plus module par le contrle dopaminergique nigro stri.
L'hypertonie parkinsonienne (rigidit) est plastique
Le tremblement parkinsonien a pour caractristique essentielle sa survenue exclusive au repos lorsque une position
de relchement musculaire est maintenue dans l'immobilit depuis quelques instants. Il s'agit d'un tremblement lent
d'amplitude variable surtout net aux extrmits. Il prdomine aux membres suprieurs ou les oscillations digitales ont
permis les comparaisons classiques comme mietter du pain, rouler une cigarette etc.
Des perturbations de la statique, de la parole, de l'criture
Elles sont la consquence de cette triade fondamentale. La marche est lente, petites enjambes. Les difficults
prdominent la mise en route ou le dmarrage est parfois impossible ou marqu par un pitinement sur place, et lors de
la rencontre d'un obstacle.
La parole est rare, lente, monotone et sourde. Ailleurs l'articulation est dfectueuse, saccade ou bloque par la
rptition itrative d'un mot ou d'une syllabe (palilalie). Le trac de l'criture est lent et irrgulier. Surtout il existe une
micrographie qui s'accentue dans le cours de la ligne.
Les dystonies
Les dystonies sont caractrises par l'apparition d'un trouble du tonus lors du maintien de certaines attitudes
(position de bout en particulier) et par sa disparition, au moins au dbut de l'affection, au repos.
Le spasme de tension
Le torticolis spasmodique
Les mouvements anormaux
Les tremblements se dfinissent par des oscillations rythmiques involontaires que dcrit tout ou partie du corps
autour de sa position d'quilibre. Le tremblement essentiel tait tremblement d'attitude. Le tremblement intentionnel
apparat au cours d'excution d'un mouvement.
Les myoclonies sont des contractions volontaires et brves d'une partie, d'un ou de plusieurs muscles, avec ou sans
dplacement du segment sur lequel ce ou ces muscles agissent.
Les mouvements choriques et athtosiques correspondent des troubles des activits musculaires synergiques.
Ils sont exagrs par l'attitude et l'activit volontaire.
126
L'quilibration et l'appareil vestibulaire
L'appareil vestibulaire adresse au systme nerveux des informations sur la position et les dplacements de la tte
dans l'espace. Une participation quilibre des deux appareils vestibulaires est ncessaire leur bon fonctionnement. La
smiologie vestibulaire s'explique par une rupture de cet quilibre physiologique. Le contrle troit des dispositifs
crbelleux, 1 efficacit des rgulations de supplance partir des affrences somesthsiques et visuelles rend compte de la
capacit d'adaptation du systme nerveux aux perturbations vestibulaires.
1 Anatomie et physiologie de l'appareil vestibulaire (fig. 7 et 8)
Les rcepteurs vestibulaires
Ils sont constitus d'un pithlium de cellules cilies dont les cils, enrobs dans une masse muco-glatineuse
(cupula) plongent dans le liquide endolymphatique. L'organisation anatomique de ces rcepteurs est celle de mcano-
rcepteurs sensibles aux dformations susceptibles de s'exercer sur les cils.
Les rcepteurs utriculaires et sacculaires sont des rcepteurs de position, sensibles la pesanteur. Leur
frquence de dcharge dpend de la position de la tte dans l'espace et ils concourent aux ractions d'adaptation
statique.
Les rcepteurs des canaux semi-circulaires sont des dtecteurs d'acclration angulaire et sont stimuls par le
dplacement du courant endolymphatique. Il existe un tonus constant de ces rcepteurs. Leur frquence de repos s'acclre
pour une rotation dans un sens dtermin et devient nulle pour une rotation en sens oppos. Ils concourent aux ractions
d'quilibration.
FIG. 7 - Appareil vestibulaire : origine, nerf, noyaux.
Nerf vestibulaire et noyaux vestibulaires
Le ganglion de Scarpa situ au fond du conduit auditif interne renferme le corps cellulaire des neurones du nerf
vestibulaire. Celui-ci accol au nerf cochlaire parcourt le conduit auditif interne puis traverse l'angle ponto-crbelleux pour
gagner dans le bulbe les noyaux vestibulaires. Ils sont au nombre de quatre (suprieur, latral, mdian, infrieur) et sont
situs sous le plancher du quatrime ventricule.
Complexes et disperses, les projections des noyaux vestibulaires se font essentiellement dans trois directions : la
moelle par le biais du faisceau vestibulo-spinal qui s'articule directement avec les motoneurones alpha ; le cervelet avec
lequel l'appareil vestibulaire a des rapports privilgis puisque d'une part les mmes noyaux qui envoient au
cervelet reoivent de lui en retour, et que d'autre part cette double projection est organise somatopiquement ;
les noyaux oculo-moteurs et la substance rticule par des fibres formant une grande partie de la bandelette
longitudinale postrieure. Certaines informations vestibulaires gagnent le thalamus et le cortex.
Intgration des messages vestibulaires
Le ganglion de Scarpa situ au fond du conduit auditif interne renferme le corps cellulaire des neurones du nerf
vestibulaire. Celui-ci accol au nerf cochlaire parcourt le conduit auditif interne puis traverse l'angle ponto-crbelleux pour
gagner dans le bulbe les noyaux vestibulaires. Ils sont au nombre de quatre (suprieur, latral, mdian, infrieur) et sont
situs sous le plancher du quatrime ventricule.
Complexes et disperses, les projections des noyaux vestibulaires se font essentiellement dans trois directions : la
moelle par le biais du faisceau vestibulo-spinal qui s'articule directement avec les motoneurones alpha ; le cervelet avec
lequel l'appareil vestibulaire a des rapports privilgis puisque d'une part les mmes noyaux qui envoient au
cervelet reoivent de lui en retour, et que d'autre part cette double projection est organise somatopiquement ;
les noyaux oculo-moteurs et la substance rticule par des fibres formant une grande partie de la bandelette
longitudinale postrieure. Certaines informations vestibulaires gagnent le thalamus et le cortex.
L'action des noyaux vestibulaires sur la musculature spinale s'exerce par l'intermdiaire de la substance
rticule et par la voie vestibulo-spinale. Les affrences vestibulaires contribuent favoriser le tonus des muscles
extenseurs et interviennent dans les rflexes de redressement. En l'absence du contrle de la vue, le redressement de la
tte est un rflexe d'origine vestibulaire qui prcde la mise en jeu des rflexes proprioceptifs assurant le redressement du
corps par rapport la tte.
Les ractions oculaires d'origine vestibulaire sont de deux types : d'une part, des ractions statiques prennent
naissance dans le systme des rcepteurs utriculaires et fixent les yeux dans une position dtermine pour chaque position
de la tte ; d'autre part des ractions phasiques prennent naissance dans les canaux semi-circulaires et sont telles que la
127
stimulation d'un canal tend provoquer un dplacement du regard dans la direction oppose. Cette dviation phasique est
lente, elle tend tre brusquement interrompue par une secousse rapide de direction oppose. La rptition de ces
secousses lentes et rapides constitue le nystagmus.
Les projections vestibulaires ascendantes qui s'intgrent au niveau de la rgion tectale, de la rgion
thalamo-pallidale et thalamo-strie, et enfin du cortex crbral, participent l'laboration du schma corporel et au contrle
du geste. Ces affrences contribuent aussi au maintien de la station debout et aux ractions d'quilibration, notamment
lorsque les affrences visuelles font dfaut. D'une faon gnrale, si les ractions la pousse sont avant tout d'ordre
proprioceptif musculaire, les ractions de l'organisme plac sur un sol mouvant sont principalement d'origine vestibulaire.
2 Smiologie vestibulaire
Le syndrome vestibulaire clinique
Le vertige labyrinthique est une sensation de rotation dans un sens dtermin donnant l'impression de
dplacement des objets environnants. La station debout et la marche peuvent tre impossibles. Des nauses, des
vomissements, divers troubles vgtatifs (sueurs, bradycardie) peuvent accompagner la sensation vertigineuse. Entrent
aussi dans le cadre des vertiges, des illusions diverses de dplacement des objets par rapport au sujet : oscillations du
cadre environnant, impression de roulis ou de dplacement vertical. Par contre les impressions vertigineuses frquemment
rencontres (sensation interne de dsquilibre, instabilit) doivent tre interprtes avec prcaution ; seuls parfois
l'exprience clinique et les examens complmentaires permettront de distinguer celles relevant d'une cause organique et
celles rencontres chez les anxieux, hystrophobiques ou dprims.
Le dsquilibre d'origine vestibulaire se manifeste chez le sujet debout, pieds joints, les yeux ferms, par
une dviation latralise de l'axe du corps qui peut aller jusqu' la chute (signe de Romberg labyrinthique). On modifie le
sens de cette dviation en changeant la position de la tte. Par ailleurs si le sujet, les yeux ferms fait successivement cinq
pas en avant, cinq pas en arrire, il apparat une dviation de la marche en direction du dficit vestibulaire (marche en
toile).
Fig. 8 - Reprsentation schmatique des noyaux vestibulaires et de leurs principales projections.
Les quatre noyaux vestibulaires sont connects entre eux et avec leurs homologues du ct oppos. Ils sont en relation
troite avec la bandelette longitudinale postrieure, le cervelet (lobe floculo-nodulare), la rticule du tronc crbral. Par
ailleurs, les informations vestibulaires gagnent le thalamus et le cortex crbral o elles se projettent au voisinage de la face
dans l'aire somato-sensible contro-latrale. Les noyaux vestibulaires exercent un contrle direct sur la moelle (faisceau
vestibulo-spinal issu du noyau de Deilers).
Les dviations segmentaires font apparatre l'action qu'exerc chaque canal semi-circulaire sur le tonus
musculaire dans son plan de fonction : dviation de l'index l'preuve des bras tendus se produisant du ct o le
labyrinthe est dficitaire.
Le nystagimis est une oscillation involontaire rythmique et conjugue des globes oculaires. Il se dfinit par le
sens dans lequel il bat, qui est, par convention, celui de la secousse rapide et par sa direction (nystagmus horizontal,
vertical, rotatoire, multidirectionnel). Le nystagmus rsultant d'une lsion priphrique est de direction mixte, horizontale et
128
rotatoire ; en revanche le nystagmus des lsions centrales apparat souvent pur, tant schmatique-ment horizontal pour
les lsions protubrantielles, rotatoire pour les lsions bulbaires et vertical pour les lsions pdonculaires.
L'exploration vestibulaire instrumentale
Elle consiste dans l'tude des ractions provoques, nystagmus surtout, lors d'une stimulation artificielle des
labyrinthes.
Dans l'preuve calorique, l'eau froide produit normalement une dviation de la tte et des yeux vers le ct
stimul et un nystagmus battant vers le ct oppos. L'eau chaude produit des effets inverses. Pour explorer les canaux
horizontaux, la tte est penche de 60 en arrire ; pour explorer les canaux verticaux, la tte est penche de 45
latralement.
L'preuve giratoire stimule les canaux semi-circulaires par rotation ; le fauteuil qui tourne la vitesse de 5
10 tours en 20 secondes est arrt brutalement. On observe la raction labyrinthique qui se produit l'arrt : le nystagmus
bat dans le sens oppos la rotation.
L'lectronystagmographie. Son principe repose sur l'enregistrement du champ rsultant de la diffrence de
potentiel qui existe entre la rtine et la corne. L'axe lectrique qui en rsulte correspond sensiblement l'axe du globe
oculaire. L'enregistrement par des lectrodes priorbitaires du champ lectrique permet d'observer fidlement les
dplacements de l'il dans les diverses directions. Les diverses varits de nystagmus spontan et provoqu peuvent tre
ainsi enregistres et quantifies.
Les deux grands types de syndrome vestibulaire
Le syndrome vestibulaire priphrique {oreille interne, nerf vestibulaire) est global, unilatral et
harmonieux, c'est--dire que les troubles sont systmatiss et se font tous dans le mme sens. Le caractre rotatoire du
vertige est net ; l'atteinte cochlaire concomitante est habituelle (hypoacousie).
Le syndrome vestibulaire central (noyaux et voies vestibulaires supra-nuclaires) est dissoci, discordant,
dysharmonieux. Il n'y a pas de latralisation franche des signes et le nystagmus est souvent marqu, l'inverse des
vertiges. Il n'y a pas d'atteinte cochlaire, mais on peut trouver d'autres signes d'atteinte du systme nerveux central.
Le contrle crbelleux
Plac en drivation sur les dispositifs sensori-moteurs le cervelet n'est indispensable aucune des formes de la
motilit mais fonctionne comme un organe de contrle extrmement labor dont la dfaillance se marque dans chacune
des activits motrices.
1 Nature et mcanisme du contrle crbelleux
Le cervelet a un rle essentiel dans le contrle du tonus et du mouvement. Il intervient dans la coordination
spatiale et temporelle des contractions musculaires lmentaires qui sous-tendent les rflexes de redressement, le maintien
de la posture rige, la modulation de l'attitude et assure la mesure (force, direction du geste propositionnel.)
L'organisation anatomique et celle de ses projections expliquent son intervention globale dans la mise en jeu de ces
diverses rgulations. Il parat tabli (v. Premire partie, chapitre III) que le cervelet mdian assure une action prdominante
sur la musculature axiale alors que les hmisphres crbelleux sont troitement lis l'laboration du geste fin et prcis
des extrmits.
Certaines donnes exprimentales suggrent une interprtation de la physiologie du cervelet en fonction de
l'quilibre entre les activits alpha et gamma. En fait l'action excitatrice des noyaux du cervelet module par le cortex
crbelleux s'exerce sur les deux types de dispositifs qui travaillent en synergie et non en concurrence.
Le cervelet dispose d'un grand nombre de boucles rtro-actives lui permettant de corriger l'activit motrice en
fonction de l'erreur dtecte suivant le principe du contrle rtro-actif (feed-back). A ce mcanisme ncessairement
imparfait semble se juxtaposer un contrle programm en fonction des acquisitions antrieures. En effet, le nombre
considrable de liaisons synaptiques au sein du cervelet suggre une fixation possible de l'exprience sous forme de
modifications structurales synaptiques. Suivant une telle conception l'automatisation d'un mouvement correspondrait la
superposition au contrle rtro-actif d'un contrle programm dont le cervelet, parfaitement inform sur l'tat du
mouvement et sur le niveau des voies effectrices, assurerait la charge en fonction des programmes imprims dans son
cortex la faveur des expriences antrieures.
2 Les manifestations fondamentales:du dficit crbelleux
L'utilisation des termes consacrs par l'usage facilite l'analyse des dsorganisations de la station debout, de la
marche, du mouvement volontaire que l'on observe chez les crbelleux. Si ces diffrentes facettes de la smiologie
crbelleuse se recoupent parfois elles n'en constituent pas moins les aspects lmentaires de la perturbation motrice
crbelleuse.
L'hypotonie musculaire donne lieu une exagration de la passivit sans hyperextensibilit des
articulations. Dans l'tude du ballant, on provoque des mouvements passifs alternatifs rapides, tirant successivement
agonistes et antagonistes d'une mme articulation. Ces preuves (ballant du poignet, ballant des paules recherch
par des mouvements imposs de rotation du tronc, ballant du pied) doivent tre juges comparativement d'un ct
par rapport l'autre.
Le caractre pendulaire des rflexes montre que cette passivit est lie la mise en jeu inefficace des antagonistes.
Dans la manuvre de Stewart-Holmes, on demande au sujet d'effectuer une contraction musculaire contre rsistance (plier
l'avant-bras sur le bras, maintenir le bras tendu en avant) puis on fait cesser brusquement la rsistance : le sujet normal
est capable de mettre fin immdiatement l'activit musculaire en cours ; le crbelleux ne peut le faire et il en rsulte un
dplacement d'autant plus ample et d'autant plus nergique que la rsistance initiale tait plus forte (la main vient par
exemple frapper l'paule).
La dysmtrie et l'hypermtrie traduisent l'amplitude exagre des mouvements (intensit et dure) chez le
crbelleux. Elles se manifestent nettement dans le geste volontaire : en demandant au malade de porter l'index sur le nez
ou le lobule de l'oreille, ou de porter le talon sur le genou du ct oppos. Que les yeux soient ouverts ou ferms, la
direction du mouvement est bonne mais son amplitude est exagre. Le mouvement dpasse son but, ce qui ncessite une
correction, elle-mme excessive.
129
L'asynergie se dfinit comme un trouble de l'association des mouvements lmentaires. Le droulement
harmonieux de l'activit motrice ncessite chaque instant une rpartition dfinie de l'activit entre diffrents muscles
synergiques et antagonistes.
Les troubles de cette harmonisation spatiale et temporelle des composantes du mouvement (dcomposition du
mouvement) constituent l'asynergie.
La dyschronomtrie est un trouble de la rapidit du mouvement. Elle dsigne aussi bien un retard anormal la
mise en route de l'activit qu'une prolongation excessive de l'action en cours. Elle apparat dans des preuves telles que faire
porter simultanment les index sur le nez.
L'adiadococinsie dsigne l'incapacit d'effectuer successivement et un rythme rapide des mouvements de
sens oppos (faire les marionnettes par exemple). L'adiadococinsie crbelleuse se marque par l'amplitude anormale des
mouvements et par la mise en jeu des territoires proximaux s'opposant la fixation posturale ncessaire une excution
normale.
Le syndrome crbelleux dans les principales formes de l'activit motrice
Dans la station debout, le trouble de l'quilibre entrane exceptionnellement la chute et se manifeste par une
instabilit anormale de l'axe du corps et des rponses rflexes d'quilibration prsentes, mais mal coordonnes et
partiellement inefficaces. La smiologie est surtout nette dans les syndromes crbelleux bilatraux, o le sujet, les bras en
abduction excessive largit son polygone de sustentation. L'quilibre antro-postrieur est imparfaitement maintenu par une
activit incessante et anarchique des jambiers antrieurs (danse des jambiers antrieurs). Les ractions la pousse sont
trop tardives pour tre efficaces et l'quilibre ne peut tre maintenu que par un brusque pas en arrire ou de ct suivant le
sens de la pousse. La mauvaise rpartition du tonus en fonction de la verticalit peut apparatre, mme en position assise ;
c'est l'ataxie du tronc.
Les perturbations de la marche, lorsqu'elles sont modres, n'apparaissent que de faon intermittente :
instabilit au demi tour, difficult la descente des escaliers, impossibilit de courir, de s'arrter brusquement ou de
contourner un obstacle. La dmarche caractristique des syndromes crbelleux est festinante : chaque pas entrane une
dviation latrale excessive et il est impossible au malade de suivre une ligne droite matrialise (dmarche pseudo-
brieuse). La mise en route est hsitante et retarde, l'arrt incertain, les pas ont une amplitude et un rythme variable.
Dans les syndromes unilatraux, la jambe du ct atteint est lance trop haut et retombe violemment, le malade avance en
zig-zag dvi du ct ls.
Le drglement des adaptations posturales est dj vident derrire les perturbations de l'quilibration et de
la marche. Il contribue aussi aux anomalies du mouvement volontaire. Nanmoins quelques tests la mettent spcialement
en vidence : lors de l'accroupissement, il se produit normalement un dcollement du talon. Ce phnomne ne se produit
pas, de faon unie ou bilatrale chez les crbelleux ; si l'on demande un sujet debout de se pencher volontairement en
arrire, on observe spontanment un certain degr de flexion des genoux. Cette adaptation posturale est ici abolie. Enfin
dans le passage de la position couche la position assise, il se produit normalement une activation des muscles du plan
postrieur des deux membres infrieurs qui les maintient sur le plan du lit. Chez le crbelleux ce phnomne ne se produit
pas et le, ou les membres infrieurs s'lvent au-dessus du plan du lit.
Parmi les perturbations des gestes propositionnels
la maladresse est parfois vidente : le malade lche involontairement les objets ; ses gestes excessifs et mal
mesurs l'exposent se heurter, renverser, casser ; il prouve de grandes difficults manipuler des objets de petite
dimension et coordonner efficacement ses mouvements lorsqu'il doit se boutonner ou remonter sa montre.
En dpit de leur caractre artificiel, les preuves usuelles (doigt-nez ; talon-genou ; renversement des mains)
montrent bien que l'asynergie, la dysmtrie, l'adiadococinsie et la dyschronomtrie sont les troubles fondamentaux qui
sous-tendent cette maladresse gestuelle.
Dans l'criture, les lettres sont de grande taille et anarchiques dans leurs dimensions ; on relve des jambages
dmesurs tmoignant de la dysmtrie. Un test simple consiste faire tracer rapidement les barreaux d'une chelle (le trait
horizontal franchit les limites verticales).
Le tremblement crbelleux n'est pas un vrai tremblement mais le rsultat d'une srie d'arrts et de
reprises dans la contraction musculaire du fait de l'asynergie, de l'asynchronisme des contractions entre muscles agonistes
et antagonistes. Il n'apparat que lors du mouvement, s'accentuant souvent du dbut la fin du geste. Il est parfois associ
un tremblement oculaire, fait d'oscillations gales, et par l diffrent d'un nystagmus. En fait ce tremblement
n'appartient qu' certaines varits de syndromes crbelleux et notamment ceux qui intressent le noyau dentel et le
pdoncule crbelleux suprieur.
La dysarthrie crbelleuse ne s'observe que dans les lsions bilatrales. La parole crbelleuse est tranante,
mal articule et en outre scande, explosive en raison d'irrgularits, non dans le rythme de l'mission, mais dans la force
de la voix qui est sans cesse changeante, d'un mot l'autre, voire d'un phonme l'autre au cours d'un mme mot.
DOCUMENTS ANNEXES extraits de "Science et Vie"
1-Comment le cerveau commande le bon geste.
Entre les gestes rflexes et les gestes intentionnels, l'tre humain dispose de la fascinante capacit d'effectuer le geste le plus
adapt son environnement. On en sait un peu plus sur la faon dont la prparation du geste, dans le cerveau, intgre les informations issues
de l'environnement, pour que les muscles effecteurs ralisent, au doigt et l'il, le bon travail.
Imaginez un instant que vous vous apprtiez raliser un geste aussi simple que prendre un verre d'eau. Essayez maintenant de
dfinir tous les dtails de l'activation et de l'inactivation des muscles impliqus. Non seulement il est difficile de nommer tous ces muscles,
mais si l'on y parvient, il sera galement difficile de dterminer l'ordre et la dure prcise de leur activation et de leur inactivation. Pourtant,
lorsque vous voyez le verre et que vous voulez le saisir pour boire, vous faites le geste sans avoir rflchir aux mcanismes utiliss par
votre cerveau, votre moelle et vos muscles. Votre cerveau dit : bouge et les muscles le font. Dans la moelle, les motoneurones contrlent
les muscles en leur commandant de se contracter dans un ordre prcis, avec la dure et la force ncessaires. Mais qu'est-ce qui contrle ces
neurones ? Le cortex crbral et plus prcisment le cortex moteur a des projections directes vers la moelle.
Le contexte fait laction
Ces petites scnes imagines montrent comment le simple stimulus (un simple verre d'eau) vous imposent des ractions bien
diffrentes : le contexte modifie l'action. C'est dire que le cerveau n'est pas une simple machine commandes. Depuis l'entre jusqu' la
sortie, tous les traitements qu'il effectue sont modifis par le contexte. Les interactions entre influences du contexte et automatismes
reprsentent une des questions les plus fascinantes de la recherche sur le contrle moteur.
On pourrait multiplier les exemples de la faon dont le systme nerveux central contrle et parfois mme limine des rflexes. Une
chose est claire : le contrle moteur est plus qu'une simple commande go . De temps en temps, une intervention consciente est ncessaire
130
pour effectuer un mouvement correct ; d'autres moments elle drange plutt. Parfois, le mouvement n'est qu'un pur rflexe ; parfois un
contrle conscient modifie ce rflexe.
Le plus lmentaire des mouvements se prsente ainsi comme une structure biomcanique d'une extrme sophistication. Devant la
ncessaire complexit du systme de contrle neuro-musculaire qui dtermine les paramtres du mouvement, l'ide s'impose que l'excution
motrice est le rsultat d'une construction labore. Cette dernire se rconciliait travers une srie d'oprations menes par le systme
nerveux central. Ces oprations permettraient de dfinir un cahier des charges dtailles, de prciser le contenu de la planification, de traduire
ce plan en un exemple structur d'ordres adresss aux muscles et, enfin, de contrler le travail de ces derniers. Le mouvement apparat donc
comme le rsultat d'une organisation strictement dtermine.
une telle construction, un fait s'oppose pourtant immdiatement : la remarquable adaptabilit de l'activit motrice. L'intervention
puissante de mcanismes de rgulation s'impose donc comme une absolue ncessit. Ces mcanismes doivent tre capables tout moment
de faire en sorte que le mouvement excut soit aussi conforme que possible la demande de l'environnement. Avant mme que
l'autorgulation sensorielle contribue aux corrections ultimes au cours du mouvement, un ensemble d'ajustements prparatoires va intervenir.
Il va modifier, affiner et dj adapter les oprations les plus prcoces de l'organisation motrice en fonction des informations disponibles.
Donc, les messages de commande mis par le cortex moteur sont eux-mmes dclenchs par d'autres messages (figure). Ces
signaux arrivent au cortex moteur en provenance d'autres aires corticales, mais aussi de structures sous corticales, comme les ganglions de
la base ou le cervelet. Ces structures envoient alors des messages au cortex moteur via une structure de relais galement sous corticale : le
thalamus. Comment ces diffrents signaux corticaux et sous-corticaux collaborent-ils pour laborer les messages envoys par le cortex
moteur vers la moelle pinire et, de l, vers les muscles ?
En 1875, le neurologue J. H Jackson proposa une organisation hirarchique du cerveau en trois niveaux. Dans cette organisation,
le cortex moteur, avec son accs direct la moelle pinire, ne se situe qu' un niveau moyen : il est soumis aux influences des rgions du
cerveau d'ordres suprieurs. Ce concept est aujourd'hui largement accept. De nombreuses observations modernes viennent en effet
renforcer l'ide d'une position intermdiaire du cortex moteur, bien qu'une simple organisation fonctionnelle via marche du cerveau soit, de
nos jours conteste. l'poque de Jackson, ces hypothses sur l'implication du cerveau dans la gense d'un acte moteur ne faisaient pas
l'unanimit chez les scientifiques.
Deux opinions diamtralement opposes ont ensuite vu le jour au dbut du xxe sicle. Pour le neurologue D. Ferrier et ses
collgues le cortex moteur tait le seul endroit responsable de l'organisation-initiation du mouvement volontaire. En revanche pour Sherrington
et ses collgues ces aires corticales n'taient que le point de dpart et non le centre du mouvement. Jackson l'avait dj prdit sur la base
dobservations de patients pileptiques ayant une pathologie crbrale.
La perception et interprtation.
L'tude de la motricit resta longtemps tributaire de contraintes mthodologiques et techniques qui limitaient l'exploration
fonctionnelle des structures nerveuses centrales. Puis le dveloppement de techniques d'enregistrement de l'activit biolectrique des cellules
nerveuses pendant la prparation et l'excution du mouvement a permis d'tudier en dtail les processus de centraux. Dans les annes 60,
le neurophysiologiste Evans fut le premier utiliser ces techniques. La question qui se posait est la suivante : lorsqu'un changement rapide
dans notre environnement exige d'entreprendre une action particulire, comment le systme nerveux central organise-t-il cette action pour
produire un comportement adapt ?
Les travaux de psychologie cognitive ont permis d'identifier trois oprations de traitements, successivement menes ds la
perception et linterprtation du signal. La premire est de slectionner, dans un rpertoire de rponse possible celle qui est adapte la
situation. Cette rponse dtermine de faon globale est symbolique. Elle ne dfinit pas un effecteur particulier, mais plutt un but
comportementale, une cible atteindre. La deuxime opration consiste dfinir les caractristiques de la rponse slectionne. Au cours de
cette opration de planification ou de programmation du mouvement est dfinie une squence de contraction musculaire ncessaire pour
raliser la rponse choisie. La troisime opration consiste excuter le mouvement. C'est elle qui est l'origine de l'activation des circuits de
commande de la musculature, responsable de la mcanique observable du geste.
Pour illustrer la squence entre la slection de la rponse et la planification du mouvement, prenons l'exemple de la saisie du verre
d'eau. Il s'agit donc de la slection de la rponse o rien n'est jusqu'alors spcifi. Mais connaissant ou non la fragilit du verre, sachant ou
non que l'eau est bouillante, voyant ou non arriver un enfant qui risque de sen saisir, vous effectuerez le mouvement trs diffremment. La
distinction entre la premire et la seconde oprations, c'est--dire entre la slection et la planification du mouvement, a dj t voqu au
sicle dernier par Jackson.
Cet auteur avait une classification des mouvements selon leur degr d'automaticit. Il dcrit un patient qui ne pouvait bouger ni la
langue ni le visage lorsqu'on le lui demandait, mais tait capable de le faire en mangeant. Il tait donc incapable d'excuter intentionnellement
un mouvement qu'il pouvait trs bien raliser de faon automatique. Ces diffrentes catgories, des plus automatiques au plus volontaires,
refltent une volution des centres nerveux vers un niveau de complexit de plus en plus grand.
Le temps de raction
L'acte moteur prsente donc deux proprits fondamentales. D'un ct, il est le rsultat d'une planification labore. De l'autre, il est
l'objet d'une rgulation permanente afin de rester conforme la demande de l'environnement. Les recherches menes en psychologie
cognitive et en neurosciences s'efforcent d'lucider cette apparente contradiction. Pour l'essentiel, on utilise un protocole exprimental dans
lequel on demande de rpondre un stimulus sensoriel par l'excution d'un mouvement particulier, puis en notant le temps qui s'coule entre
l'apparition de ce stimulus et le dbut du mouvement. On appelle ce temps coul le temps de raction . Ce type d'expriences permet de
tirer des conclusions sur le temps minimum ncessaire pour dclencher une activit motrice en rponse une simulation sensorielle.
Le raisonnement est simple. Il repose sur l'hypothse que ce temps de raction est celui qu'il faut au systme nerveux pour
effectuer les oprations de traitements depuis le stimulus jusqu' l'initiation du mouvement demand. On peut alors observer comment ce
temps varie en fonction de l'information, fournie par avance au sujet, sur les caractristiques du mouvement. Le temps de raction est d'autant
plus court que le sujet dispose d'informations pralables. Il est modifi non seulement par la quantit de l'information, mais aussi par sa
nature. La conclusion la plus importante qu'on peut tirer de ce type d'expriences et que l'information pralable doit intervenir pendant le
processus de planification du mouvement. Ce fait doit amliorer l'efficacit de ce processus et contribuer ainsi au raccourcissement du temps
de raction. La question est, ds lors, de savoir quels mcanismes neuronaux sont impliqus et quel rle joue le cortex moteur dans ce
processus.
Afin de mieux comprendre ce qui se passe, on peut tudier les diffrentes tapes de la construction d'un acte moteur de manire
slective. Pour ce faire, on analyse les changements d'activit lectrique des neurones individuels du cortex moteur chez un singe conditionn
effectuer une tache motrice. Le protocole de la tche utilise dans ces tudes suit toujours le mme principe. Lors d'un essai exprimental,
deux signaux sont prsents successivement. Le signal informatif, prsent en premier, fournit une information pralable sur une ou plusieurs
caractristiques du mouvement que l'animal doit effectuer aprs l'apparition d'un deuxime signal, le signal impratif. On peut conditionner
des singes une preuve sensori-motrice simple : par exemple, pointer droite et ou gauche sur une cible lumineuse plus ou moins
loigne. Le signal informatif fournit soit une information complte (l'animal connat d'avance, c'est--dire avant l'arrive du signal impratif, la
direction et l'amplitude du mouvement) soit une information partielle (l'animal connat seulement davance la direction ou lamplitude). Les
effets obtenus sont trs clairs : le temps de raction est plus court lorsque l'information pralable porte sur la direction plutt que sur
l'amplitude du mouvement.
Les enregistrements de l'activit unitaire de neurones du cortex moteur mettent en vidence trois catgories de neurones. Dans la
premire catgorie, on trouve des neurones activs seulement pendant l'excution du mouvement, donc seulement aprs l'apparition du
second signal, le signal impratif. Ceci n'est gure surprenant puisque ces neurones font partie du cortex moteur considr comme
131
responsable de la production des mouvements. Les neurones de la deuxime catgorie sont activs seulement par la prsentation du premier
signal, le signal informatif, et ceci, trs souvent, slectivement par rapport au contenu informationnel de ce signal. Aucune trace d'activit lie
au mouvement ne peut tre dcele. C'est dj beaucoup plus surprenant : cela montre en effet que le cortex moteur n'est pas purement
moteur , mais galement associatif . Enfin les neurones de la troisime catgorie prsentent simultanment les deux proprits, reliant
donc un processus un autre.
Ces rsultats mettent en lumire deux points particulirement intressants. Tout d'abord, le dcours temporel de l'activit des trois
catgories de neurones correspond clairement au droulement du processus ncessaire la production du mouvement. Ensuite, l'activit
slective des diffrents neurones montre leur implication respective dans les oprations spcifiques diffrents paramtres du mouvement,
comme la direction, l'amplitude, la force, etc.
Ceci explique, en partie, comment le systme nerveux central assume son rle, face une stimulation : slectionner une rponse
comportementale, programmer le mouvement adquat et, enfin, l'excuter. Cependant, ceci n'explique pas encore ce qui dtermine le
raccourcissement du temps de raction. On a pu montrer que l'activit d'une cellule nerveuse augmente la suite d'un stimulus informatif. On
a pu montrer que le temps de raction est plus court lorsqu'on dispose d'une information pralable, par rapport la situation o elle fait dfaut.
Il parat donc logique de suggrer que cette activit neuronale slective est, d'une faon ou d'une autre, l'origine de ce raccourcissement du
temps de raction.
Un test trs simple consiste rechercher une corrlation statistique entre le taux d'activation neuronale et le temps de raction. Le
rsultat de ce test montre qu'il existe en effet une corrlation entre ces deux facteurs. Il suggre donc que l'augmentation de l'activit
neuronale, en fonction de l'information fournie par le premier stimulus, induit un raccourcissement du temps de raction.
Transformation en srie
Envisags tant du point de vue de la psychologie cognitive que des neurosciences, les mcanismes qui assurent la transition entre
le traitement de l'information apporte par le signal et l'excution d'une rponse motrice adapte soulve une question cruciale si l'on veut
comprendre la remarquable adaptabilit comportementale dont font preuve les mammifres suprieurs. les rponses qu'y ont apportes les
psychologues exprimentaux comme les neurophysiologistes ont longtemps fait appel la seule notion de fonctionnement sriel .
Cela signifie que le rsultat de la transformation de l'information effectue au cours d'une opration de traitements serait ensuite
transmis une autre opration, o il ferait l'objet d'une autre transformation plus labore. La question qui se pose est donc de savoir si les
oprations de traitements des informations, depuis la perception du stimulus jusqu' l'excution du mouvement, se droulent en stricte
succession (une opration ne pouvant commencer avant que la prcdente soit termine) ou si elles peuvent tre menes simultanment.
Une exprimentation simple permet d'lucider cette question. Un singe doit soit flchir, soit le poignet la suite d'une discrimination
perceptible trs simple, donc rapidement accomplie, entre deux signaux lumineux, l'une prsente sa droite l'autre sa gauche. Cependant,
l'excution du mouvement va, en plus, dpendre d'une discrimination perceptible difficile, donc lentement accomplie, entre deux stimulus
situs trs prs l'un de l'autre. Bref, selon l'emplacement prcis du stimulus, le mouvement doit tre soit excut, soit retenu. Question : le
systme moteur va-t-il commencer programmer la rponse sur la base premire information qui lui est transmise ? celle concernant donc la
direction du mouvement du premier, flexion extension, et attendre l'instruction lui indiquant de l'excuter ou non, ou alors entreprendra-t-il
cette programmation qu'une fois en possession de ces deux informations complmentaires ?
l'analyse de l'activit neuronale du cortex moteur confirme l'hypothse d'un traitement parallle . Une partie des neurones
impliqus dans la spcification de la direction du mouvement est active ds que celle-ci est connue, ce, alors mme que le systme perceptif
n'a pas encore communiqu l'information ncessaire pour prendre la dcision d'excuter ou non le mouvement.
Jusqu'ici, nous avons seulement parl de neurones individuels et de leur contribution diffrents processus. Cependant, tout le
monde sait aujourd'hui que le traitement de l'information, mme le plus lmentaire, n'est pas reprsent dans l'activit d'un seul neurone,
mais dans l'activit globales de centaines voire de milliers de neurones. De nombreux neurones du cortex crbral changent dactivit en
fonction des diffrents paramtres du mouvement, par exemple la direction. Cependant, on ne sait pas trs bien comment, en tant que
membre d'un groupe, ils interagissent seuls maxillaires et collaborent lors de l'laboration d'une tache motrice.
Comment la direction du mouvement tait le ct au sein des groupes de neurones du cortex moteur ? l'exprience suivante
apporte les lments de rponse. D'abord, on entrane un singe excutif du mouvement de pointage, de qu'une cible centrale, dans toutes les
directions distribues alentour. Lorsqu'on enregistre pendant l'excution de cette tche l'activit des neurones du cortex moteur, on peut
constater que beaucoup d'entre eux module leur activit en fonction de la direction du mouvement. Mais contrairement ce qu'on pensait
jusqu'alors, le spectre de leur slectivit n'est pas trs troit, mais plutt large. En effet bien que chaque neurone prsente une dcharge
maximale pour une direction donne, sa direction prfre et, il reste nanmoins actif au cours de mouvements orients diffremment. Il
est d'autant plus que la trajectoire de mouvement se rapproche davantage de son orientation prfre. Donc, la notion d'une large
comptence (ou, ce qui revient au mme, d'une faible spcialisation) nous loigne de la mtaphore du clavier de moteur cortical. Les
touches , dont chacune correspondrait un muscle, ce serait activ en squence pour composer la mlodie cintique des commandes
neuromusculaires. Mais cette ide que la participation d'un neurone la spcification directionnelle du mouvement est plus quantitative que
qualitative une consquence plus importante encore : la spcification du mouvement doit ncessairement faire intervenir une collaboration
fonctionnelle entre neurones.
Lorsqu'on fait l'addition aprs un calcul appropri, de l'activit de toute une population de neurones, on constate que la slectivit de
la population est beaucoup plus prcise que celles d'un neurone individuel. La figure quatre montre en effet que la direction du mouvement
(un mouvement vers la cible numro trois) et reprsent dans l'activit de la population de faon prcise, et ceci, mme pendant la priode
prparatoire, donc entre l'apparition des deux signaux, pendant laquelle l'animale devait en fait tre immobile.
L'tude de la collaboration et de l'interaction entre neurones faisant partie des populations entires et donc une tche
particulirement fascinante qui va nous aider mieux comprendre la gense de l'laboration d'un acte moteur et donc du comportement.
2-Les premiers pas.
Comme pour toute activit motrice complexe, l'acquisition de la marche implique la maturation du systme nerveux. Mais pas
seulement. Pour l'enfant, elle correspond aussi, et peut-tre avant tout, un vritable apprentissage.
Chaque parent a pu observer la succession des stages qui accompagnent le dveloppement postural et moteur de l'enfant : d'abord
la matrise de la tte, suivie de celle de la position assise, puis de la position debout, enfin de la marche. Gesell, en 1946, a expliqu cette
succession par un dveloppement orient et progressif. Orient : du haut du corps jusqu'au bas du corps (dveloppement cphalo-caudal) et
pour chaque membre des segments les plus proches de l'axe du corps au plus loigns (proximo-distal). Progressif : chaque tape impliquait
un accroissement du contrle du tonus musculaire et de la coordination des mouvements rendus possibles par la maturation du systme
nerveux. Un aspect de ce dveloppement continue d'intriguer les parents comme les spcialistes. Quel lien y a-t-il entre la marche
automatique du nourrisson est celle que l'enfant commence matriser vers un an ? la ressemblance entre ces deux activits est en effet
frappante. l'instar de ce qu'il fera plus tard tout seul, le nouveau-n, maintenu en position verticale et en appui sur une surface plane, produit
un mouvement altern des jambes. L'explication la plus souvent avance reprenait les thses de McGraw (1943). La marche du nouveau-n
est une activit rflexe contrle par des centres sous-corticaux. Quand, avec le dveloppement du contrle cortical, se met en place une
activit motrice de mieux en mieux matrise, cette activit rflexe est inhibe.
Un courant de pense plus rcent aborde la question d'une manire diffrente. Thelen (1983) observa que cette activit persiste
dans le pdalage que produit l'enfant de quatre six semaines couch sur le dos ou maintenu immerg mi-corps en position verticale.
l'oppos, si l'on ajoute une masse sa jambe ( quatre semaines la masse ajoute est gale au poids moyen gagn dans les deux semaines
132
suivantes), la frquence de mouvement de cette dernire diminue. De mme, cinq ou six mois, priode laquelle on n'observe
gnralement aucune activit alterne des jambes, celle-ci apparat quand l'enfant est maintenue soulev sur un tapis roulant. Il adapte mme
sa frquence de pas la vitesse de droulement du tapis (Thelen, 1991). Que montrent ces travaux ? Qu'en manipulant les conditions
d'exercice d'une activit alterne des jambes qui rappelle celle de la marche, on peut soit la provoquer, soit la rduire.
Pour effectuer cette activit locomotrice (marche, course, saut cloche-pied, ou toute activit impliquant le corps tout entier) de
nombreux systmes sont mobiles. On peut en citer quelques-uns : le systme musculaire qui permet le dplacement des segments corporels
ainsi que les ajustements posturaux ; les oscillateurs spinaux qui contrlent l'activit rythmique alterne des membres (Grillner, 1975) ; les
systmes sensoriels, visuel, labyrinthique et proprioceptif ; le systme physique en action, c'est--dire le corps dont les caractristiques
anthropomtriques varient avec l'ge ; le systme cognitif charg de traiter les informations disponibles, etc. on peut donc considrer toute
activit motrice comme la rsultante d'une activit intgre de ces diffrents systmes. Ces caractristiques dpendront ainsi du mode de
fonctionnement de chacun des systmes mobiliss et des conditions contextuelles de leur fonctionnement selon les proprits de la tche
effectuer.
L'activit locomotrice d'un enfant qui, vers 12 mois, commence marcher, est en fait assez diffrente de celle de l'adulte. Les
caractristiques des premiers pas de marche indpendante sont bien connues. Les pas sont petits. L'cartement des pieds est important. La
frquence de pas est plus grande que chez l'adulte. La marche ne s'accompagne pas d'un balancement des bras. Le pos du pied se fait
plat et non par le talon. La marche en canard est frquente. D'un point de vue musculaire, on observe de nombreuses contractions des
muscles agonistes et antagonistes.
Comme mode de locomotion humaine, la marche consiste dplacer son corps selon un axe antro-postrieur, grce une
succession de simple-appui (appui unipodal) et de double-appuis (appui bipodal). Ce dplacement, au plan fonctionnel, la caractristique
essentielle de la tche locomotrice, ne sera possible que grce la cration de force propulsives. Ces dernires sont gnres par la cration
et la modulation de deux distances entre eux le centre de pression (le barycentre des forces ractives au sol) et la projection au sol du centre
de gravit du marcheur.
Cette activit conduite une situation de dsquilibre. De fait, le centre de pression et le centre de gravit du sujet ne se trouve
peu prs jamais sur la mme verticale (dfinition de l'quilibre : pour qu'un corps rigide reposant sur un plan horizontal soit en quilibre, deux
conditions doivent tre runies : un, la rsultante des forces externes, la raction R du plan et des forces gravitaires, mg, poids du corps, doit
tre gal zro ; deux, la somme des moments de toutes ces forces relativement G, centre de gravit, doit tre nulle. Une consquence de
ces deux conditions est une distance nulle entre le barycentre des forces ractives, centre de pression, et la projection au sol du centre de
gravit. En outre, si la projection du centre de gravit se trouve un intrieur de la surface d'appui dlimit par le contour des deux pieds, le
polygone de sustentation, alors l'quilibre peut tre rtabli. Il n'en est pas de mme si le centre de gravit se projette l'extrieur du polygone
de sustentation, situation qui entrane alors une chute de l'objet considr). Toutefois, au cours d'un cycle, c'est--dire entre deux contacts au
sol du mme pied, la distance si qui sparent ces deux points sera plus ou moins grande. En outre, la projection du centre de gravit peut se
faire hors de la base de sustentation. C'est ce qui se passe durant la phase d'appui unipodal o le dsquilibre est alors maximal. On
comprend ainsi l'importance d'une activit posturale constante pour contrebalancer la stabilit lie une activit mme de la marche et viter
une perte d'quilibre conduisant ventuellement une chute.
Pour l'enfant, les dbuts de la marche reprsentent une exprience totalement nouvelle : il lui faudra tre capable de produire et de
contrler la succession de dsquilibres unipodaux et bipodaux ncessaires une progression dans le plan antro-postrieur. Or cette
poque, l'enfant est incapable de matriser l'quilibre sur un pied, il n'acquerra cette capacit que plus tard. Apprendre marcher peut ainsi
tre dcrit comme le processus d'intgration de ncessits posturales et dynamiques. Les premires portent sur la stabilisation du corps pour
viter une perte d'quilibre est donc une chute ; les secondes, sur la cration des conditions dynamiques permettant de dplacer le corps vers
l'avant. Il faut par consquent apprendre intgrer, c'est--dire interprter et utiliser les informations sensorielles disponibles.
Les rsultats d'une tude longitudinale suggrent que l'acquisition de la marche autonome par l'enfant est un processus en deux
tapes.
La premire phase, interprte comme une phase d'intgration de la posture et du mouvement, dure, selon les enfants, de trois
cinq mois. Elle est caractrise par une volution trs rapide des paramtres de la marche. Passant de 0,20 m/s 0,80 m/s, la vitesse
moyenne de progression augmente, ainsi que la longueur du pas et la fourchette de vitesses utilises ; la marche devient vritablement
fonctionnelle. Simultanment, l'cartement des pieds diminue fortement ainsi que la dure relative du double appui. Ces deux derniers
paramtres sont gnralement considrs comme indexant les aspects posturaux de la marche. L'enfant qui commence marcher utilise une
base de support trs large, comparativement l'adulte. Il minimise ainsi les consquences possibles du dsquilibre produit. De mme,
l'importante dure du double appui est interprte comme ncessaire au bon rtablissement de l'quilibre. On observe paralllement la mise
en place d'une corrlation significative entre frquence de pas et vitesse comme c'est le cas chez l'adulte. D'o l'hypothse suivante : durant
cette premire phase d'acquisition de la marche, afin de rsoudre un probable moteur particulier, dplacer son corps vers l'avant par une
succession d'appui uni et bipodaux, l'enfance apprend utiliser les contraintes mcaniques du systme compos de son corps, du sol sur
lequel il marche et de la gravit.
Pour autant, le fait que la marche devienne efficace n'implique pas que l'enfant utilise la mme stratgie de propulsion que
l'adulte chez l'un et chez l'autre, les valeurs de l'acclration verticale au cours du pas apparaissent en effet trs diffrentes. Cette
acclration est interprte comme un indicateur de l'activit antigravitaire. En d'autres termes, elle renseigne sur la capacit maintenir la
position du centre de gravit un niveau qui oscille de manire priodique autour d'une constante pour une succession de pas. l'extrme fin
du simple appui, cette valeur, positive chez l'adulte, et toujours ngative chez l'enfant, ce qui traduit une situation de chute dynamique du
centre de gravit.
Une longue exprience.
La seconde phase est une phase d'ajustement. Elle correspond une priode d'affinement du contrle de la marche. C'est durant
cette tape, au terme d'environ six sept annes de marche indpendante, c'est--dire partir de l'ge de sept ou huit ans, que l'enfant
parvienne produire une stratgie de propulsion de type adulte.
Cette priode est marque par une volution caractristique des valeurs de l'acclration verticale mesure l'instant de la
transition entre le simple appui et le double appui, moments de dsquilibre maximal. Aprs trois annes de marche indpendante, ces
valeurs deviennent positives (les valeurs adultes ne seront cependant atteintes environ que deux ans plus tard) : la stratgie de propulsion a
chang l'enfant n'est plus en situation de chute la fin du simple appui. Il est devenu capable, grce un contrle posturale unipodal plus
efficace, de se propulser durant le simple appui. Ces rsultats vont dans le sens des tudes qui ont montr que, avant l'ge de sept ans,
l'enfant n'utilise pas dans la marche un pattern d'activit musculaire semblable celui de l'adulte.
C'est aussi durant cette priode que semblent se dvelopper les phnomnes anticipateurs (spatiaux et temporels) de la marche
tels qu'ils ont t dcrits chez l'adulte. Une analyse de la phase d'initiation de la marche chez des enfants commenant marcher, ainsi que
chez des enfants de 2,5 8 ans, permet de faire l'hypothse que la capacit d'un contrle pro actif de l'action n'existe pas au dpart mais se
construit progressivement avec l'exprience locomotrice. Ce contrle pro actif du mouvement recouvre deux fonctions essentielles : 1) il
permet d'anticiper le dsquilibre produit par le passage d'un appui bipodal un un appui unipodal, et donc de s'y prparer ; 2) l'activit
anticipatrice initie les conditions de l'action venir. Ce deuxime aspect a t notre connaissance moins systmatiquement explor. Sa
fonction dans le cas de la marche est trs explicite : cette activit anticipatrice conditionne la cration de forces propulsives. Lors de la phase
de transition entre la position debout stable et la premire oscillation de la jambe, une des caractristiques du comportement anticipateur tient
en un recul du centre des pressions (barycentre des forces ractives au sol) avant que ce dernier ne se dplace dans le sens de la marche.
Ce recul est une valeur absolue proportionnel la vitesse qui sera atteinte la fin du premier pas. Il sert donc crer des conditions
dynamiques de la marche.
133
134
Le mouvement de l'enfant qui commence marcher ne prsente aucune des caractristiques de l'anticipation. Ce n'est qu'aprs
quelques mois de marche, c'est--dire au dbut de cette deuxime phase d'acquisition, que l'on en observera certaines caractristiques de
manire plus ou moins alatoire tout d'abord puis plus systmatiquement quatre ans, c'est--dire aprs environ trois annes d'exprience de
la marche. La prsence de comportements anticipateurs ne signifie toutefois pas que ceux-ci soient organiss comme ils le sont chez l'adulte.
En particulier, ce recul du centre des pressions n'est corrl avec la vitesse venir qu' partir de 6 ans environ. Vers 8 ans, les valeurs de la
corrlation restent encore nettement plus faibles que chez l'adulte. On constate donc que l'acquisition d'un contrle nature de la marche
ncessite pour l'enfant de sept huit annes d'exprience !
Comme cela a dj t dit, le jeune marcheur pose le pied plat. Chez l'adulte, au contraire le contact se fait par le talon. Une
analyse de la frquence de contact par le talon avec l'ge de marche montre que ce comportement apparat brusquement au moment de la
transition entre les deux priodes mises en vidence.
Comment expliquer ce dveloppement prsent chez chaque enfant ? Si l'apparition du contact par le talon rsultait directement de
la maturation des neurones comme cela est suggr, on ne voit pas pourquoi il apparatrait pour chaque enfant la fin de la premire priode
d'apprentissage de la marche et ce, quelle que soit la dure de cette priode. Pour certains, l'absence de contact par le talon serait due l'tat
immature du systme nerveux de l'enfant que, qui ne permettrait pas la dorsiflexion du pied durant l'oscillation de la jambe. Or, bien avant le
dbut de la marche autonome, on peut observer cette dorsiflexion qui est le plus souvent suivi dun pos de pied plat ! Notre hypothse est
que ce comportement est d manque de matrise posturale. Les caractristiques du mouvement du tout jeune marcheur (pas grands, grand
cartement de pieds, dure relative du double appui importante, etc.) sont ici interprtes comme une adaptation fonctionnelle la stabilit de
l'enfant ne permettant pas le contact par le talon. Ce ne serait qu' partir du moment o l'enfant a acquis une matrise posturale suffisante que
se mettrait en place un mode de locomotion plus efficace impliquant l'utilisation du contact par le talon.
La ncessaire exploration de l'espace.
En conclusion nous aimerions souligner l'intrt et la ncessit de distinguer, dans les tudes sur le mouvement, diffrents niveaux
de causalit, chacun relevant de logiques explicatives diffrentes. Dans les activits locomotrices, le corps se dplace mon espace grce la
cration de forces qui engendrent un mouvement. un autre niveau, il est ncessaire de considrer l'activit neuro-musculaire indispensable
pour engendrer ces forces, diffrents patterns d'activit musculaire pouvant conduire, un niveau global, au mme rsultat du point de vue
des forces gnres. C'est pourquoi une explication de l'acquisition de la marche en termes essentiellement de maturation du systme
nerveux, plus largement du systme neuro-musculaire, nous semble partielle. Si un certain degr de maturation du systme nerveux est
indispensable la mise en place d'activits motrices complexes, un telle que la marche, il n'en explique pas directement les caractristiques
comportementales.
On insiste sur le fait que l'acquisition de la marche correspondrait pour l'enfant un vritable apprentissage. On peut en effet poser
hypothse que la mise en place d'un contrle fin de la marche ncessite l'exploration de l'espace de la tche. C'est elle qui permettrait la
construction, dveloppement, de stratgies locomotrices adaptes l'tat du systme locomoteur et l'environnement immdiat.
Dans cette perspective il devient plus ais d'expliquer pourquoi cet enfant de 14 mois, bon marcheur, se remet soudain quatre
pattes lorsqu'il se trouve face une pente de 15 20. Le changement d'environnement a en effet entran une modification importante de la
tche. Monter ou plus encore, descendre une pente ncessite pour l'enfant de construire une nouvelle stratgie de production et de
modulation de forces propulsives trs diffrentes l'une de l'autre. La mise en place de cette nouvelle stratgie implique que l'enfant puisse
explorer les proprits du nouveau couplage environnement-tche et dcouvrir de nouvelles solutions motrices adaptes.
La situation de l'enfant non voyant est elle aussi intressante. Des rsultats prliminaires ne suggreraient pas de diffrence notable
dans le stratgie motrice utilise par ces enfants dans les premires semaines de marche. Ainsi ce ne serait qu'au moment o l'enfant a la
capacit de se dplacer aisment dans l'espace, que l'absence de vision le contraindrait adopter une stratgie qui lui vite de heurter un
obstacle. L'enfant utilise une stratgie d'exploration de l'espace partir d'une succession de pas dont un sur deux a une longueur nulle (aprs
avoir fait un pas, le pied controlatral est ramen hauteur de l'autre). Il a t not que pour lun des enfants il suffisait que la mre mette la
main sur son paule, lui procurant ainsi un point d'ancrage dans l'espace, pour qu'il adopte une stratgie de marche semblable l'enfant
voyant. Il a donc suffi qu'un lment nouveau, apparemment mineur, mais essentiel dans le cas de l'enfant non voyant, pour que l'organisation
comportementale de son action soit substantiellement modifie.
Vous aimerez peut-être aussi
- Musculation Des BradsDocument20 pagesMusculation Des Bradsderdouri63% (8)
- La Puissance Du RegardDocument13 pagesLa Puissance Du RegardSaravorn100% (11)
- Rtoubles Vesico-SphincteriensDocument106 pagesRtoubles Vesico-SphincteriensFares KhalfallahPas encore d'évaluation
- Techniques - Reeducatives - Pelvi Périnéologie - Sportive - Hau - Niveau PDFDocument36 pagesTechniques - Reeducatives - Pelvi Périnéologie - Sportive - Hau - Niveau PDFMjidKarimPas encore d'évaluation
- N Syndrome Pyramidal 2015Document24 pagesN Syndrome Pyramidal 2015richardPas encore d'évaluation
- Sémiologie NeuroDocument10 pagesSémiologie Neuroimma_drPas encore d'évaluation
- Le Sang Et La Circulation SanguineDocument1 pageLe Sang Et La Circulation SanguineAgnès FromentPas encore d'évaluation
- Le Guide Pratique Des Etirements ReussisDocument31 pagesLe Guide Pratique Des Etirements Reussisdiourou100% (1)
- Décrypter Le Langage Du Corps PDFDocument68 pagesDécrypter Le Langage Du Corps PDFAmeziane Youcef OualiPas encore d'évaluation
- Le Système Nerveux VégétatifDocument15 pagesLe Système Nerveux Végétatifcoursenligne.lille1Pas encore d'évaluation
- Hygiène de la voix et gymnastique des organes vocaux: Histoire de la musique depuis son origine jusqu'à nos joursD'EverandHygiène de la voix et gymnastique des organes vocaux: Histoire de la musique depuis son origine jusqu'à nos joursPas encore d'évaluation
- PCEM1 2008 Cours 1bis OsDocument10 pagesPCEM1 2008 Cours 1bis Osphilippefr0% (1)
- Systeme Nerveux Autonome PDFDocument28 pagesSysteme Nerveux Autonome PDFMina HappinessPas encore d'évaluation
- Procréation: Les Grands Articles d'UniversalisD'EverandProcréation: Les Grands Articles d'UniversalisPas encore d'évaluation
- Contraction MusculaireDocument2 pagesContraction Musculairemohbio_869948847100% (1)
- QCM MuscleDocument2 pagesQCM Musclele sagePas encore d'évaluation
- Physiologie Musculaire 2020Document47 pagesPhysiologie Musculaire 2020drahlemtrifPas encore d'évaluation
- 6.les Troubles de La SensibilitéDocument6 pages6.les Troubles de La SensibilitéHøpë FûllPas encore d'évaluation
- Neurophy 2Document203 pagesNeurophy 2Adel KchoukPas encore d'évaluation
- Neurophy 2Document203 pagesNeurophy 2Adel KchoukPas encore d'évaluation
- Anatomie SNC Texte PDFDocument95 pagesAnatomie SNC Texte PDFJean PatatePas encore d'évaluation
- Cours de Reeducation VestibulaireDocument106 pagesCours de Reeducation VestibulaireZOHEIR RAMDANE100% (1)
- 10-09-07 - Les Chaines MusculairesDocument7 pages10-09-07 - Les Chaines MusculairescharbelhannaPas encore d'évaluation
- II Les Réflexes ProprioceptifsDocument31 pagesII Les Réflexes ProprioceptifsHamza Abu Humaid100% (2)
- Diagnostic Neuropathies Peripheriques RecommandationsDocument28 pagesDiagnostic Neuropathies Peripheriques RecommandationsHakimoPas encore d'évaluation
- EcoleDocument7 pagesEcoleWilfried Hess MalotPas encore d'évaluation
- Eval Clinique SpasticitéDocument9 pagesEval Clinique SpasticitéJean Patate0% (1)
- Neuroanatomie Complet JustineDocument123 pagesNeuroanatomie Complet JustineMaria OlteanPas encore d'évaluation
- Physiologie Neuro-MuscuclaireDocument53 pagesPhysiologie Neuro-MuscuclaireMampionona RakotonandrasanaPas encore d'évaluation
- La Contraction MusculaireDocument11 pagesLa Contraction MusculaireJihadi AbbouPas encore d'évaluation
- Entraînement neuro-athlétique pour débutants Plus de coordination, de mobilité et de concentration grâce à l'amélioration de la neuro-athlétisation - plan de 10 semaines inclusD'EverandEntraînement neuro-athlétique pour débutants Plus de coordination, de mobilité et de concentration grâce à l'amélioration de la neuro-athlétisation - plan de 10 semaines inclusPas encore d'évaluation
- ContractionDocument13 pagesContractionPY00Pas encore d'évaluation
- TD Reflexe MyotatiqueDocument4 pagesTD Reflexe MyotatiquelemiaPas encore d'évaluation
- Système Nerveux AutonomeDocument21 pagesSystème Nerveux AutonomeArtichaudPas encore d'évaluation
- 01-Somesthésie 12Document17 pages01-Somesthésie 12nourhoudaPas encore d'évaluation
- $le Tonus Musculaire Et Ses TroublesDocument53 pages$le Tonus Musculaire Et Ses Troublesmehdi.chlif4374Pas encore d'évaluation
- 2.introduction À La Sémiologie de La MotricitéDocument1 page2.introduction À La Sémiologie de La MotricitérichardPas encore d'évaluation
- Le Systeme Nerveux AutonomeDocument5 pagesLe Systeme Nerveux AutonomesafemindPas encore d'évaluation
- Acte Réflexe Intéressant CompletDocument4 pagesActe Réflexe Intéressant CompletFagarou rekPas encore d'évaluation
- 5.les Troubles de La Fonction MotriceDocument14 pages5.les Troubles de La Fonction MotriceHøpë FûllPas encore d'évaluation
- Nerfs CraniensDocument2 pagesNerfs CraniensIdrissou Fmsb100% (1)
- ProprioceptionDocument3 pagesProprioceptionUSJHANDBALLPas encore d'évaluation
- Cours Histologie Du Cervelet 1Document11 pagesCours Histologie Du Cervelet 1Dany MelPas encore d'évaluation
- 7 Cervelet PDFDocument8 pages7 Cervelet PDFŽĭņëb BėñPas encore d'évaluation
- 3 SomesthésieDocument25 pages3 SomesthésieSahouin Claude MedardPas encore d'évaluation
- CerveletDocument7 pagesCerveletDeaƭh SurgeơnPas encore d'évaluation
- Neurologie - Cours N°19 - Rousseau - Syndrome FrontalDocument18 pagesNeurologie - Cours N°19 - Rousseau - Syndrome FrontalCoralie PascardPas encore d'évaluation
- Examen de La Marche Et de La Station DeboutDocument3 pagesExamen de La Marche Et de La Station DeboutAbdellatif OutrahPas encore d'évaluation
- Muscle Et Physiologie MusculaireDocument20 pagesMuscle Et Physiologie MusculaireBoubker PouperPas encore d'évaluation
- Neuroanat-2 0Document83 pagesNeuroanat-2 0seloua kaid100% (1)
- Physiologie Cardio Vasculaire 3Document32 pagesPhysiologie Cardio Vasculaire 3Faïza SahraouiPas encore d'évaluation
- LombalgieDocument29 pagesLombalgieSkyIsTheLimit BăăPas encore d'évaluation
- La Reeducation Pbve Fevrier08Document66 pagesLa Reeducation Pbve Fevrier08Alexis GoncalvesPas encore d'évaluation
- MOSTA Physiologie de La Jonction NeuromusculaireDocument27 pagesMOSTA Physiologie de La Jonction NeuromusculaireSamia BelabbasPas encore d'évaluation
- La Fonction RespiratoireDocument143 pagesLa Fonction RespiratoireRoxana RoxPas encore d'évaluation
- Vieilissement MusculaireDocument66 pagesVieilissement Musculaireghinsavit0% (1)
- SD CerebDocument22 pagesSD Cereb31243564Pas encore d'évaluation
- Parasympathomemetiques Et ParasympatholytiquesDocument46 pagesParasympathomemetiques Et ParasympatholytiquesAMINA KHOUDRIAPas encore d'évaluation
- L-Appareil Locomoteur 1Document41 pagesL-Appareil Locomoteur 1Trou BailPas encore d'évaluation
- These MezouriDocument105 pagesThese MezouriichkhuyPas encore d'évaluation
- Biomecanique Et Analyse de La MarcheDocument64 pagesBiomecanique Et Analyse de La Marcheti tiPas encore d'évaluation
- 2-Systeme MusculaireDocument6 pages2-Systeme MusculaireahlemmPas encore d'évaluation
- Synapse Et Transmission SynaptiqueDocument10 pagesSynapse Et Transmission SynaptiqueFaculté De Médecine BécharPas encore d'évaluation
- Le Muscle PDFDocument121 pagesLe Muscle PDFYounes BroukiPas encore d'évaluation
- Anat Palpa EpauleDocument5 pagesAnat Palpa EpauleSolaine Saint-LouisPas encore d'évaluation
- Résumé G.Chakib NerveuxDocument5 pagesRésumé G.Chakib NerveuxMaroua AbzPas encore d'évaluation
- Tissu Musculaire 1Document92 pagesTissu Musculaire 1Adonis Serghini100% (2)
- Physiologie Noyaux Gris Centraux PidouxDocument10 pagesPhysiologie Noyaux Gris Centraux PidouxOwassiPas encore d'évaluation
- 2 S7 Cours 11Document29 pages2 S7 Cours 11Chabane OubarechePas encore d'évaluation
- anatomieIII PDFDocument10 pagesanatomieIII PDFSoufiane Sassi100% (1)
- Médecine Personnalisée: Utilisation du profil génétique pour le traitement de la maladieD'EverandMédecine Personnalisée: Utilisation du profil génétique pour le traitement de la maladiePas encore d'évaluation
- Habermas - L'Espace Publique 30 Ans AprèsDocument32 pagesHabermas - L'Espace Publique 30 Ans AprèsLuis Boris SantosPas encore d'évaluation
- CG2 TH3 Mod3 IntroAnaMouv 20131030Document28 pagesCG2 TH3 Mod3 IntroAnaMouv 20131030charbelhannaPas encore d'évaluation
- Enonce EthiqueDocument16 pagesEnonce Ethiquecharbelhanna0% (1)
- Habermas PDFDocument562 pagesHabermas PDFcharbelhannaPas encore d'évaluation
- 0301 Modalit S Mobilisations APDocument8 pages0301 Modalit S Mobilisations APcharbelhannaPas encore d'évaluation
- La Trompe D'EustacheDocument5 pagesLa Trompe D'EustachecharbelhannaPas encore d'évaluation
- Bilan MorphostatiqueDocument69 pagesBilan MorphostatiquecharbelhannaPas encore d'évaluation
- 0301 Cheville Pied - Mvts A P - Goniom TrieDocument9 pages0301 Cheville Pied - Mvts A P - Goniom Triecharbelhanna100% (1)
- 10 Bilans Articulaires Cliniques Et Goniometriques GeneralitesDocument8 pages10 Bilans Articulaires Cliniques Et Goniometriques GeneralitescharbelhannaPas encore d'évaluation
- Ob ChainesDocument13 pagesOb ChainescharbelhannaPas encore d'évaluation
- Chaines Fonctionnelles MSDocument50 pagesChaines Fonctionnelles MSSimona Alice Onea100% (1)
- 04 - Techniques de Kinï¿ Sithï¿ Rapie ActiveDocument4 pages04 - Techniques de Kinï¿ Sithï¿ Rapie ActiveJedidi AmirPas encore d'évaluation
- DocumentDocument137 pagesDocumentcharbelhannaPas encore d'évaluation
- Fiche 06 - Travail ActifDocument4 pagesFiche 06 - Travail ActifcharbelhannaPas encore d'évaluation
- 04 - Techniques de Kinï¿ Sithï¿ Rapie ActiveDocument4 pages04 - Techniques de Kinï¿ Sithï¿ Rapie ActiveJedidi AmirPas encore d'évaluation
- 04 - Techniques de Kinï¿ Sithï¿ Rapie ActiveDocument4 pages04 - Techniques de Kinï¿ Sithï¿ Rapie ActiveJedidi AmirPas encore d'évaluation
- Grandes Fon CtionDocument23 pagesGrandes Fon CtioncharbelhannaPas encore d'évaluation
- Cours 3 EMG - 2Document99 pagesCours 3 EMG - 2ismailPas encore d'évaluation
- Mi08 15Document63 pagesMi08 15geantdounePas encore d'évaluation
- Physiologie Et Biochimie Du Muscle Alimentaire 2Document10 pagesPhysiologie Et Biochimie Du Muscle Alimentaire 2Zacharie Carime BassaPas encore d'évaluation
- 10 Façons de Libérer Les Pouvoirs Du Nerf VagueDocument12 pages10 Façons de Libérer Les Pouvoirs Du Nerf VagueMoncef MejriPas encore d'évaluation
- Qe 2017Document11 pagesQe 2017Ahmed Baichit0% (1)
- Corrige: Sciences de La Vie Et de La Terre GroupeDocument4 pagesCorrige: Sciences de La Vie Et de La Terre GroupekathyPas encore d'évaluation
- Histologie Et Embryologie MedicalesDocument200 pagesHistologie Et Embryologie MedicalesLakezis100% (2)
- Integration Par Les Neurones Exercices CorrigesDocument3 pagesIntegration Par Les Neurones Exercices CorrigesMamadou Kane100% (1)
- Connaissances G2n2rales en Zootechnie 2016Document164 pagesConnaissances G2n2rales en Zootechnie 2016chimène ollePas encore d'évaluation
- PlyometricDocument7 pagesPlyometriclatifaPas encore d'évaluation
- Le Suivi Therapeutique PharmacologiqueDocument134 pagesLe Suivi Therapeutique PharmacologiqueazizaPas encore d'évaluation
- These HuaCAODocument169 pagesThese HuaCAOJean-Serein MbendePas encore d'évaluation
- gp09009 Delaere Trigger 1ere Partie PDFDocument4 pagesgp09009 Delaere Trigger 1ere Partie PDFIulian IvasciucPas encore d'évaluation
- SVT Obli G1Document8 pagesSVT Obli G1LETUDIANTPas encore d'évaluation
- Etude Corrélative Entre Les Deux Mouvements Olympiques Et Le SquatDocument146 pagesEtude Corrélative Entre Les Deux Mouvements Olympiques Et Le SquatAmin GasmiPas encore d'évaluation
- 621f06c7dc573-BAC - SVT - Série DDocument16 pages621f06c7dc573-BAC - SVT - Série DaxelPas encore d'évaluation
- BIO 270 Physiologie Cellulaire Générale 2020 PDFDocument125 pagesBIO 270 Physiologie Cellulaire Générale 2020 PDFTONDI ABDOU CHACOURPas encore d'évaluation
- CM Musculation L2 2017 PDFDocument118 pagesCM Musculation L2 2017 PDFYAELKALYSSE100% (1)
- Préparation À l'ET Musculation Exemples de QuestionsDocument2 pagesPréparation À l'ET Musculation Exemples de QuestionsAbdelkarim ElaliPas encore d'évaluation
- 125 Codage Nerveux 76Document20 pages125 Codage Nerveux 76gheorgheadrianoneaPas encore d'évaluation
- 21svtj2me1 Corrige DefDocument14 pages21svtj2me1 Corrige DefmeninoPas encore d'évaluation
- Comment Le Muscle Strié Squelettique TransformeDocument19 pagesComment Le Muscle Strié Squelettique TransformeBentouina mohammed saidPas encore d'évaluation