Vous aimerez peut-être aussi
- Plant Cell Wall Patterning and Cell ShapeD'EverandPlant Cell Wall Patterning and Cell ShapeHiroo FukudaPas encore d'évaluation
- NIH Public Access: Bacterial Swarming: A Model System For Studying Dynamic Self-AssemblyDocument31 pagesNIH Public Access: Bacterial Swarming: A Model System For Studying Dynamic Self-AssemblyverdosPas encore d'évaluation
- Reviews: Bacterial Biofilms: From The Natural Environment To Infectious DiseasesDocument15 pagesReviews: Bacterial Biofilms: From The Natural Environment To Infectious DiseasesJordy KunPas encore d'évaluation
- General Microbiology (Chapter 3)Document18 pagesGeneral Microbiology (Chapter 3)Ashraf OsmanPas encore d'évaluation
- Biofilms: Microbial Life On Surfaces: Rodney M. DonlanDocument10 pagesBiofilms: Microbial Life On Surfaces: Rodney M. DonlanKapil JhajhariaPas encore d'évaluation
- Cell Locomotion PDFDocument8 pagesCell Locomotion PDFmanoj_rkl_07Pas encore d'évaluation
- Traffic - 2004 - Etienne Manneville - Actin and Microtubules in Cell Motility Which One Is in ControlDocument8 pagesTraffic - 2004 - Etienne Manneville - Actin and Microtubules in Cell Motility Which One Is in ControlPinaki NayakPas encore d'évaluation
- Can Bacteria WalkDocument6 pagesCan Bacteria WalkInternational Journal of Innovative Science and Research TechnologyPas encore d'évaluation
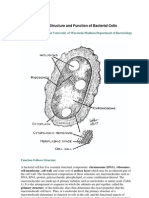
- Morphology and Fine StructureDocument67 pagesMorphology and Fine StructureDiksha PadamPas encore d'évaluation
- Bab 6 Sruktur Dan Fungsi Sel Bakteria (I)Document9 pagesBab 6 Sruktur Dan Fungsi Sel Bakteria (I)Shakira AmiraPas encore d'évaluation
- 2013 Chapter IntechDocument27 pages2013 Chapter IntechMiguel Fernandez NiñoPas encore d'évaluation
- AsifaDocument46 pagesAsifaPGMBslidesPas encore d'évaluation
- Opposite and Coordinated Rotation of Amphitrichous Flagella Governs Oriented Swimming and Reversals in A Magnetotactic SpirillumDocument8 pagesOpposite and Coordinated Rotation of Amphitrichous Flagella Governs Oriented Swimming and Reversals in A Magnetotactic SpirillumFernandoPas encore d'évaluation
- Autophagy and Disease: Always Two Sides To A Problem: Invited ReviewDocument19 pagesAutophagy and Disease: Always Two Sides To A Problem: Invited ReviewCLPHtheoryPas encore d'évaluation
- Bacterial FlagellaDocument15 pagesBacterial FlagellaSergio D. PérezPas encore d'évaluation
- Tanyut Huidrom - Assignment 3 - PhysiologyDocument5 pagesTanyut Huidrom - Assignment 3 - PhysiologyTanyut HuidromPas encore d'évaluation
- Bio FilmDocument7 pagesBio FilmFREIMUZICPas encore d'évaluation
- Microbial Anatomy - Prokaryotic MicrobesDocument55 pagesMicrobial Anatomy - Prokaryotic MicrobesRoel Jason TuboPas encore d'évaluation
- FLGV Forms A Flagellar Motor Ring That Is Required For Optimal Motility of Heliobacter PyloriDocument18 pagesFLGV Forms A Flagellar Motor Ring That Is Required For Optimal Motility of Heliobacter PyloriChris WeberPas encore d'évaluation
- Cell Biology of Hyphae: Oded YardenDocument8 pagesCell Biology of Hyphae: Oded YardenAngus YanPas encore d'évaluation
- Introduction To The Histology and Cell Biology of The Actin CytoskeletonDocument2 pagesIntroduction To The Histology and Cell Biology of The Actin CytoskeletonpajtgocjPas encore d'évaluation
- The Physical Chemistry of Cytoplasm and Its Influence On Cell Function: An UpdateDocument4 pagesThe Physical Chemistry of Cytoplasm and Its Influence On Cell Function: An UpdateIvan Jason SalyanoPas encore d'évaluation
- Allama Iqbal Open University IslamabadDocument24 pagesAllama Iqbal Open University IslamabadAbdullah NagraPas encore d'évaluation
- Chapter 7-Membrane Structure and Function : Life at The EdgeDocument5 pagesChapter 7-Membrane Structure and Function : Life at The EdgeMurry PumaPas encore d'évaluation
- Mathematical Models of Cell Motility: &) J. P. Mcgarry P. E. MchughDocument15 pagesMathematical Models of Cell Motility: &) J. P. Mcgarry P. E. MchughVishesh ManglaPas encore d'évaluation
- Living Liquid Crystals: Shuang Zhou, Andrey Sokolov, Oleg D Lavrentovich, Igor S AransonDocument32 pagesLiving Liquid Crystals: Shuang Zhou, Andrey Sokolov, Oleg D Lavrentovich, Igor S AransonTyler Jordan DavisonPas encore d'évaluation
- Analyzing Circulating Tumor Cells One at A Time: HighlightsDocument22 pagesAnalyzing Circulating Tumor Cells One at A Time: Highlightsian merisPas encore d'évaluation
- Reviews: Mechanomicrobiology: How Bacteria Sense and Respond To ForcesDocument14 pagesReviews: Mechanomicrobiology: How Bacteria Sense and Respond To ForcesOded SabahPas encore d'évaluation
- BIO350 BacStrucDocument65 pagesBIO350 BacStrucPrincewill SeiyefaPas encore d'évaluation
- Galley Proof Ms Ijbpas 2021 5610Document9 pagesGalley Proof Ms Ijbpas 2021 5610saisree malladiPas encore d'évaluation
- Blue and White Professional Science Project PresentationDocument26 pagesBlue and White Professional Science Project PresentationAbdul Matin RahimiPas encore d'évaluation
- The Bacterial FlagellumDocument1 pageThe Bacterial Flagellummferra100% (1)
- Tmp4e28 TMPDocument11 pagesTmp4e28 TMPFrontiersPas encore d'évaluation
- Engineering Biofilm Formation and Dispersal: Thomas K. Wood, Seok Hoon Hong and Qun MaDocument8 pagesEngineering Biofilm Formation and Dispersal: Thomas K. Wood, Seok Hoon Hong and Qun MaShampa SenPas encore d'évaluation
- Ultrastructure of Bacteria & EndosporeDocument8 pagesUltrastructure of Bacteria & EndosporeSubrata KunduPas encore d'évaluation
- Cytoskeleton - 2012 - Farr - Cytokinesis in TrypanosomesDocument11 pagesCytoskeleton - 2012 - Farr - Cytokinesis in TrypanosomesDario DiosPas encore d'évaluation
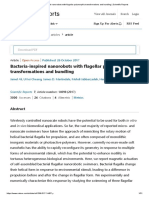
- Bacteria-Inspired Nanorobots With Flagellar Polymorphic Transformations and Bundling - Scientific ReportsDocument31 pagesBacteria-Inspired Nanorobots With Flagellar Polymorphic Transformations and Bundling - Scientific ReportsZamfirMarianPas encore d'évaluation
- (15) - Swanson, J. a. Текст СтатьиDocument24 pages(15) - Swanson, J. a. Текст СтатьиDenisPas encore d'évaluation
- BBB 0007Document6 pagesBBB 0007Tecnoy ManuelPas encore d'évaluation
- 1 s2.0 S1350417713001193 MainDocument8 pages1 s2.0 S1350417713001193 MainIITRANANDJADHAVPas encore d'évaluation
- Chemo TaxisDocument18 pagesChemo TaxisJames FranklinPas encore d'évaluation
- CambioDocument16 pagesCambioEdson AugustoPas encore d'évaluation
- Content ServerDocument10 pagesContent ServerAdit VekariaPas encore d'évaluation
- Flagellar Restoration in C. ReinhardiDocument8 pagesFlagellar Restoration in C. Reinhardinasek22Pas encore d'évaluation
- Intra-Colony Motility of Microcystis Wesenbergii CellsDocument8 pagesIntra-Colony Motility of Microcystis Wesenbergii Cellssya135949Pas encore d'évaluation
- The Zipper Mechanism in Phagocytosis: Energetic Requirements and Variability in Phagocytic Cup ShapeDocument17 pagesThe Zipper Mechanism in Phagocytosis: Energetic Requirements and Variability in Phagocytic Cup ShapeshabanaPas encore d'évaluation
- Teacher: DR Gouri Course: Zoology Hons. 6 Sem Paper: Developmental BiologyDocument17 pagesTeacher: DR Gouri Course: Zoology Hons. 6 Sem Paper: Developmental BiologyApratim SinghPas encore d'évaluation
- Biofilm ReportDocument50 pagesBiofilm ReportMary Joy DalisayPas encore d'évaluation
- BacteriologyDocument11 pagesBacteriologyCarmelle Zia Reyes100% (1)
- Animal BT Assignment 1Document10 pagesAnimal BT Assignment 1Alabhya DasPas encore d'évaluation
- Assignment For Membrane Structure: Ane Austin Lynn N. Rebancos Bsed-Science 3ADocument9 pagesAssignment For Membrane Structure: Ane Austin Lynn N. Rebancos Bsed-Science 3Ajane austin lynn rebancosPas encore d'évaluation
- Membrane Potentials in Living Systems, Tools To Measure: Biological BackgroundDocument10 pagesMembrane Potentials in Living Systems, Tools To Measure: Biological BackgroundazzaassPas encore d'évaluation
- Membrane and Cell Wall Deposition at The Division SiteDocument2 pagesMembrane and Cell Wall Deposition at The Division SiteNatalia LadinoPas encore d'évaluation
- Harshada and PranavDocument13 pagesHarshada and PranavPranav NakhatePas encore d'évaluation
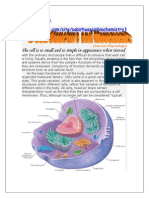
- Cell Structure by Sabir.h.nDocument15 pagesCell Structure by Sabir.h.nprinceaimy.bestPas encore d'évaluation
- Cell Surface MembraneDocument3 pagesCell Surface Membraneb111broomPas encore d'évaluation
- Science Wps OfficeDocument16 pagesScience Wps OfficeLuke Chase JacintoPas encore d'évaluation
- Gurt Flip Flop Jpcb07Document6 pagesGurt Flip Flop Jpcb07Dinok DiblongPas encore d'évaluation
- Dan Et Al-2010-Biotechnology and BioengineeringDocument10 pagesDan Et Al-2010-Biotechnology and BioengineeringNithiananthan K VejayanPas encore d'évaluation
- Cell Regulation Continually Redefining The - 1998 - Current Opinion in MicrobioDocument4 pagesCell Regulation Continually Redefining The - 1998 - Current Opinion in MicrobioJuan Pablo Vizcaya GuependoPas encore d'évaluation
- On How To Design A Low Voltage SwitchboardDocument11 pagesOn How To Design A Low Voltage SwitchboardsabeerPas encore d'évaluation
- Course Weekly Schedule Health Science TheoryDocument6 pagesCourse Weekly Schedule Health Science Theoryapi-466810096Pas encore d'évaluation
- AACO 7th Aviation Fuel Forum: AttendanceDocument3 pagesAACO 7th Aviation Fuel Forum: AttendanceJigisha Vasa0% (1)
- EE2401 Power System Operation and ControlDocument93 pagesEE2401 Power System Operation and ControlPrasanth GovindarajPas encore d'évaluation
- Method of Toxicity Test 1Document59 pagesMethod of Toxicity Test 1Widya AnggrainiPas encore d'évaluation
- The Preparation of Culture MediaDocument7 pagesThe Preparation of Culture MediaNakyanzi AngellaPas encore d'évaluation
- Educational Facility Planning: Bsarch V-2 Arch. Rey GabitanDocument20 pagesEducational Facility Planning: Bsarch V-2 Arch. Rey Gabitanidealistic03Pas encore d'évaluation
- Meng Mammogram JulDocument4 pagesMeng Mammogram JulLivia MillPas encore d'évaluation
- Dynamic Stretching - Stability - Strength.570239Document2 pagesDynamic Stretching - Stability - Strength.570239Sylvia GracePas encore d'évaluation
- Meditran SX Sae 15w 40 API CH 4Document1 pageMeditran SX Sae 15w 40 API CH 4Aam HudsonPas encore d'évaluation
- NeoResin DTM Presentation 9-01Document22 pagesNeoResin DTM Presentation 9-01idreesgisPas encore d'évaluation
- DTC P1602 Deterioration of Battery: DescriptionDocument5 pagesDTC P1602 Deterioration of Battery: DescriptionEdy SudarsonoPas encore d'évaluation
- Withania DSC PDFDocument6 pagesWithania DSC PDFEfsha KhanPas encore d'évaluation
- 365 Days (Blanka Lipińska)Document218 pages365 Days (Blanka Lipińska)rjalkiewiczPas encore d'évaluation
- Defects Lamellar TearingDocument6 pagesDefects Lamellar Tearingguru_terexPas encore d'évaluation
- B737-800 Air ConditioningDocument7 pagesB737-800 Air ConditioningReynaldoPas encore d'évaluation
- Notes, MetalsDocument7 pagesNotes, MetalsindaiPas encore d'évaluation
- Wire Rope Inspection ProgramDocument2 pagesWire Rope Inspection Programسيد جابر البعاجPas encore d'évaluation
- BDC 6566Document6 pagesBDC 6566jack.simpson.changPas encore d'évaluation
- The Human Body: An Orientation: Part ADocument10 pagesThe Human Body: An Orientation: Part ARoi Christoffer Jocson PeraltaPas encore d'évaluation
- Silly VersesDocument29 pagesSilly Verseskevin dalePas encore d'évaluation
- House of Candy PresentationDocument42 pagesHouse of Candy PresentationRohit JaroudiyaPas encore d'évaluation
- MSDS of Poly Aluminum ChlorideDocument5 pagesMSDS of Poly Aluminum ChlorideGautamPas encore d'évaluation
- Usp 1078 Good Manufacturing Practices For Bulk Pharmaceutical Excipients PDFDocument13 pagesUsp 1078 Good Manufacturing Practices For Bulk Pharmaceutical Excipients PDFMayson BaliPas encore d'évaluation
- Silage Sampling and AnalysisDocument5 pagesSilage Sampling and AnalysisG_ASantosPas encore d'évaluation
- 2015 4-H Show & Sale CatalogDocument53 pages2015 4-H Show & Sale CatalogFauquier NowPas encore d'évaluation
- Fortune Falls - Lou VaneDocument402 pagesFortune Falls - Lou VanetestePas encore d'évaluation
- Recommended Practice For Corrosion Management of Pipelines in Oil & Gas Production and TransportationDocument123 pagesRecommended Practice For Corrosion Management of Pipelines in Oil & Gas Production and Transportationsaifoa100% (2)
- 1 SMDocument10 pages1 SMAnindita GaluhPas encore d'évaluation
- Poverty and Children's Personal RelationshipsDocument87 pagesPoverty and Children's Personal RelationshipsJoe OglePas encore d'évaluation
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessD'Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessÉvaluation : 4 sur 5 étoiles4/5 (33)
- Return of the God Hypothesis: Three Scientific Discoveries That Reveal the Mind Behind the UniverseD'EverandReturn of the God Hypothesis: Three Scientific Discoveries That Reveal the Mind Behind the UniverseÉvaluation : 4.5 sur 5 étoiles4.5/5 (52)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsD'EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsÉvaluation : 4.5 sur 5 étoiles4.5/5 (6)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisD'EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisÉvaluation : 3.5 sur 5 étoiles3.5/5 (2)
- Why We Die: The New Science of Aging and the Quest for ImmortalityD'EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityÉvaluation : 4.5 sur 5 étoiles4.5/5 (6)
- Who's in Charge?: Free Will and the Science of the BrainD'EverandWho's in Charge?: Free Will and the Science of the BrainÉvaluation : 4 sur 5 étoiles4/5 (65)
- Gut: the new and revised Sunday Times bestsellerD'EverandGut: the new and revised Sunday Times bestsellerÉvaluation : 4 sur 5 étoiles4/5 (393)
- Masterminds: Genius, DNA, and the Quest to Rewrite LifeD'EverandMasterminds: Genius, DNA, and the Quest to Rewrite LifePas encore d'évaluation
- Tales from Both Sides of the Brain: A Life in NeuroscienceD'EverandTales from Both Sides of the Brain: A Life in NeuroscienceÉvaluation : 3 sur 5 étoiles3/5 (18)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedD'EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedÉvaluation : 4 sur 5 étoiles4/5 (11)
- The Rise and Fall of the Dinosaurs: A New History of a Lost WorldD'EverandThe Rise and Fall of the Dinosaurs: A New History of a Lost WorldÉvaluation : 4 sur 5 étoiles4/5 (597)
- Seven and a Half Lessons About the BrainD'EverandSeven and a Half Lessons About the BrainÉvaluation : 4 sur 5 étoiles4/5 (111)
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouD'EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouÉvaluation : 4.5 sur 5 étoiles4.5/5 (62)
- Good Without God: What a Billion Nonreligious People Do BelieveD'EverandGood Without God: What a Billion Nonreligious People Do BelieveÉvaluation : 4 sur 5 étoiles4/5 (66)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorD'EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorPas encore d'évaluation
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionD'EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionÉvaluation : 4 sur 5 étoiles4/5 (812)
- Change Your Brain, Change Your Life (Before 25): Change Your Developing Mind for Real-World SuccessD'EverandChange Your Brain, Change Your Life (Before 25): Change Your Developing Mind for Real-World SuccessÉvaluation : 4 sur 5 étoiles4/5 (18)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesD'EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesÉvaluation : 4.5 sur 5 étoiles4.5/5 (397)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceD'EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceÉvaluation : 4.5 sur 5 étoiles4.5/5 (517)
- Remnants of Ancient Life: The New Science of Old FossilsD'EverandRemnants of Ancient Life: The New Science of Old FossilsÉvaluation : 3 sur 5 étoiles3/5 (3)
- Darwin's Doubt: The Explosive Origin of Animal Life and the Case for Intelligent DesignD'EverandDarwin's Doubt: The Explosive Origin of Animal Life and the Case for Intelligent DesignÉvaluation : 4 sur 5 étoiles4/5 (19)
- Moral Tribes: Emotion, Reason, and the Gap Between Us and ThemD'EverandMoral Tribes: Emotion, Reason, and the Gap Between Us and ThemÉvaluation : 4.5 sur 5 étoiles4.5/5 (116)
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomD'EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomÉvaluation : 4 sur 5 étoiles4/5 (216)
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindD'EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindÉvaluation : 4.5 sur 5 étoiles4.5/5 (93)
- Civilized To Death: The Price of ProgressD'EverandCivilized To Death: The Price of ProgressÉvaluation : 4.5 sur 5 étoiles4.5/5 (215)