Vous aimerez peut-être aussi
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (119)
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (399)
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (587)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2219)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (344)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (890)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (73)
- MICRO (1) Fundamentals of Microbiology - Dra - AbuevaDocument16 pagesMICRO (1) Fundamentals of Microbiology - Dra - AbuevaCindy Mae Macamay100% (1)
- Antibiotic Susceptibility TestingDocument11 pagesAntibiotic Susceptibility TestingReman AlingasaPas encore d'évaluation
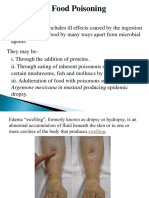
- Food Poisoning and IntoxicationsDocument26 pagesFood Poisoning and IntoxicationslakshmijayasriPas encore d'évaluation
- Vitek 2 System Principle PDFDocument5 pagesVitek 2 System Principle PDFArshia NazirPas encore d'évaluation
- Case Study of MicrobiologyDocument4 pagesCase Study of MicrobiologyQaisrani Y9Pas encore d'évaluation
- Mode of Infection of DiseaseDocument14 pagesMode of Infection of Diseasekhalid100% (1)
- Adenovirus Antigen, StoolDocument2 pagesAdenovirus Antigen, StoolNuseatPas encore d'évaluation
- Uv LightDocument5 pagesUv LightKingHungTingPas encore d'évaluation
- H5N1 FAQs: Bird Flu Questions AnsweredDocument3 pagesH5N1 FAQs: Bird Flu Questions AnsweredFiera RiandiniPas encore d'évaluation
- Peptococcus PeptostreptococcusDocument25 pagesPeptococcus PeptostreptococcusIon PopescuPas encore d'évaluation
- Skripsi KustaDocument61 pagesSkripsi Kustaandre yuindartantoPas encore d'évaluation
- Aim: Determination of RPR or VDRL RPR: Rapid Plasma Reagin VDRL: - Venereal Disease Research LaboratoryDocument3 pagesAim: Determination of RPR or VDRL RPR: Rapid Plasma Reagin VDRL: - Venereal Disease Research LaboratoryMotherterrasa VocationalPas encore d'évaluation
- Oncogenic VirusesDocument37 pagesOncogenic VirusesSalman Majid0% (1)
- Some Behavioral and Legal Considerations While Travelling To AustraliaDocument2 pagesSome Behavioral and Legal Considerations While Travelling To AustraliaRafael ValderramaPas encore d'évaluation
- Lista Clsi-2014 VTKDocument1 pageLista Clsi-2014 VTKMessy ToapantaPas encore d'évaluation
- College of Our Lady of Mt. Carmel Public Health Course SyllabusDocument4 pagesCollege of Our Lady of Mt. Carmel Public Health Course Syllabus;'SiLeNt';Pas encore d'évaluation
- STD PRESENTATIONDocument89 pagesSTD PRESENTATIONKaarthigan RamaiahPas encore d'évaluation
- Monkey Pox Fact SheetDocument3 pagesMonkey Pox Fact SheetMalcolm M. LeePas encore d'évaluation
- E2799-12 Standard Test Method For Testing DisinfecDocument9 pagesE2799-12 Standard Test Method For Testing DisinfecA MusaverPas encore d'évaluation
- Studying Microorganisms in The Living StateDocument6 pagesStudying Microorganisms in The Living StateEloisa BraillePas encore d'évaluation
- Emerging Infectious Diseases - Vol. 14Document167 pagesEmerging Infectious Diseases - Vol. 14rvcf100% (1)
- WHO SurveillanceVaccinePreventable 21 Typhoid R1Document13 pagesWHO SurveillanceVaccinePreventable 21 Typhoid R1Adre Dwi WiratamaPas encore d'évaluation
- Antibiotic Resistance Gene Testing of Recycled Water Samples in Flagstaff, AZDocument7 pagesAntibiotic Resistance Gene Testing of Recycled Water Samples in Flagstaff, AZkrtica8061Pas encore d'évaluation
- Tuberculosis Quiz: Accelerating Advocacy For TB/HIV 15th July 2010, ViennaDocument32 pagesTuberculosis Quiz: Accelerating Advocacy For TB/HIV 15th July 2010, ViennaMansoor Ali KhanPas encore d'évaluation
- New COVID-19 entry protocols for FilipinosDocument5 pagesNew COVID-19 entry protocols for FilipinosmonchanPas encore d'évaluation
- 3478-Article Text-13352-1-10-20220210Document12 pages3478-Article Text-13352-1-10-20220210Danu MuhammadPas encore d'évaluation
- Epidemiology and Public HealthDocument21 pagesEpidemiology and Public HealthFatima SumabatPas encore d'évaluation
- Test Report: Test Results Biol. Ref. Result For Sars-Cov-2 (Covid-19) Negative CT Value of Orf1Ab Gene (If Positive)Document2 pagesTest Report: Test Results Biol. Ref. Result For Sars-Cov-2 (Covid-19) Negative CT Value of Orf1Ab Gene (If Positive)Ishaan KumarPas encore d'évaluation
- SBL 1023 Exp 5Document9 pagesSBL 1023 Exp 5api-383623349Pas encore d'évaluation
- Antimicrobials in A Nutshell 27 JuneDocument37 pagesAntimicrobials in A Nutshell 27 JuneNikita NanwaniPas encore d'évaluation