Vous aimerez peut-être aussi
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (121)
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (588)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (266)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (399)
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5794)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2259)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (895)
- Olive Leaf Extract (HIV Fighter, Yeast Buster, Viral Killer)Document16 pagesOlive Leaf Extract (HIV Fighter, Yeast Buster, Viral Killer)XR500Final100% (8)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (73)
- Human Excretory SystemDocument2 pagesHuman Excretory SystemVio ElenaPas encore d'évaluation
- Hiv Associated Hematological Malignasis - Marcus HentrichDocument239 pagesHiv Associated Hematological Malignasis - Marcus HentrichJaronjer1Pas encore d'évaluation
- (José Das Neves, Bruno Sarmento (Eds.) ) Mucosal DDocument603 pages(José Das Neves, Bruno Sarmento (Eds.) ) Mucosal Dfaysal100% (1)
- Confidential 1 Pr110 WatermarkDocument51 pagesConfidential 1 Pr110 WatermarkMarissa AldePas encore d'évaluation
- Els Reviewer 2nd QuarterDocument8 pagesEls Reviewer 2nd QuarterKaezzy Yen RequermePas encore d'évaluation
- Nonmalignant Leukocyte Disorder Chapter 26-35Document15 pagesNonmalignant Leukocyte Disorder Chapter 26-35Reizel GaasPas encore d'évaluation
- Who Searo Dengue Situation Update 24 Sep 2012Document3 pagesWho Searo Dengue Situation Update 24 Sep 2012Michael KevinPas encore d'évaluation
- Field Evaluation of A Lethal Ovitrap Against Dengue Vectors in BrazilDocument7 pagesField Evaluation of A Lethal Ovitrap Against Dengue Vectors in BrazilMichael KevinPas encore d'évaluation
- Field Evaluation of A Lethal Ovitrap Against Dengue Vectors in BrazilDocument7 pagesField Evaluation of A Lethal Ovitrap Against Dengue Vectors in BrazilMichael KevinPas encore d'évaluation
- Designing of A Laboratory Model For Evaluation of The Residual Effects of Deltamethrin (K-Othrine WP 5%)Document7 pagesDesigning of A Laboratory Model For Evaluation of The Residual Effects of Deltamethrin (K-Othrine WP 5%)Michael KevinPas encore d'évaluation
- Identification of Elevated Esterase Activity in A Pyrethroid-Resistant Population of Anopheles Albimanus WiedemannDocument4 pagesIdentification of Elevated Esterase Activity in A Pyrethroid-Resistant Population of Anopheles Albimanus WiedemannMichael KevinPas encore d'évaluation
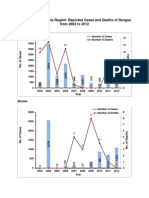
- WHO Searo Reported Cases and Deaths of Dengue From 2003 To 2012Document5 pagesWHO Searo Reported Cases and Deaths of Dengue From 2003 To 2012Michael KevinPas encore d'évaluation
- Development of Sensitive Esterase Assays Based On a-Cyano-Containing EstersDocument9 pagesDevelopment of Sensitive Esterase Assays Based On a-Cyano-Containing EstersMichael KevinPas encore d'évaluation
- Hammock MRF Proposal Progress ReportDocument4 pagesHammock MRF Proposal Progress ReportMichael KevinPas encore d'évaluation
- Potential Development of Temephos Resistance in Aedes Aegypti Related To Its Mechanism and Susceptibility To Dengue VirusDocument6 pagesPotential Development of Temephos Resistance in Aedes Aegypti Related To Its Mechanism and Susceptibility To Dengue VirusMichael KevinPas encore d'évaluation
- Mosquito Carboxylesterases A Review of The Molecular Biology and Biochemistry of A Major Insecticide Resistance MechanismDocument12 pagesMosquito Carboxylesterases A Review of The Molecular Biology and Biochemistry of A Major Insecticide Resistance MechanismMichael KevinPas encore d'évaluation
- Bioassay and Biochemical Analyses of Insecticide Resistance in Southern African Anopheles Funestus (Diptera Culicidae)Document8 pagesBioassay and Biochemical Analyses of Insecticide Resistance in Southern African Anopheles Funestus (Diptera Culicidae)Michael KevinPas encore d'évaluation
- Variation in Susceptibility Status To Organophosphate Insecticide Among Several Geographic Populations of Aedes Albopictus Skuse in IndonesiaDocument5 pagesVariation in Susceptibility Status To Organophosphate Insecticide Among Several Geographic Populations of Aedes Albopictus Skuse in IndonesiaMichael KevinPas encore d'évaluation
- Insecticide Resistance and Vector ControlDocument9 pagesInsecticide Resistance and Vector ControlMichael KevinPas encore d'évaluation
- Larva Aedes Aegypti Sudah Toleran Terhadap Temepos Di Kota Banjarbaru, Kalimantan SelatanDocument19 pagesLarva Aedes Aegypti Sudah Toleran Terhadap Temepos Di Kota Banjarbaru, Kalimantan SelatanMichael KevinPas encore d'évaluation
- Insecticide Resistance Development in CulexDocument8 pagesInsecticide Resistance Development in CulexMichael KevinPas encore d'évaluation
- Dengue Prevention and 35 Years of Vector Control in SingaporeDocument7 pagesDengue Prevention and 35 Years of Vector Control in SingaporeMichael KevinPas encore d'évaluation
- Bottle and Biochemical Assays On Temephos Resistance in Aedes Aegypti in Thailand PDFDocument9 pagesBottle and Biochemical Assays On Temephos Resistance in Aedes Aegypti in Thailand PDFMichael KevinPas encore d'évaluation
- Biochemical Detection of Pyrethroid Resistance Mechanism in AedesDocument7 pagesBiochemical Detection of Pyrethroid Resistance Mechanism in AedesMichael KevinPas encore d'évaluation
- Daftar PustakaDocument5 pagesDaftar PustakaMichael KevinPas encore d'évaluation
- Biochemical Detection of Pyrethroid Resistance Mechanism in AedesDocument7 pagesBiochemical Detection of Pyrethroid Resistance Mechanism in AedesMichael KevinPas encore d'évaluation
- Hammock MRF Proposal Progress ReportDocument4 pagesHammock MRF Proposal Progress ReportMichael KevinPas encore d'évaluation
- Current Resistance Status To Temephos in Aedes AegyptiDocument4 pagesCurrent Resistance Status To Temephos in Aedes AegyptiMichael KevinPas encore d'évaluation
- Daftar PustakaDocument5 pagesDaftar PustakaMichael KevinPas encore d'évaluation
- Progress NovDocument1 pageProgress NovMichael KevinPas encore d'évaluation
- Cell Theory - Bio 1Document41 pagesCell Theory - Bio 1Anand Wealth JavierPas encore d'évaluation
- What Is GM CornDocument2 pagesWhat Is GM CornjamilcuzzPas encore d'évaluation
- Science-8 - Q4 - WHLP 2021-2022Document8 pagesScience-8 - Q4 - WHLP 2021-2022Zerah ElsisuraPas encore d'évaluation
- Viral VaccinesDocument7 pagesViral VaccinesMoon niePas encore d'évaluation
- Drug Development ProcessapolloDocument19 pagesDrug Development Processapollotamara_0021Pas encore d'évaluation
- Effective Microbial Bioremediation Via The Multi-Omics Approach: An Overview of Trends, Problems and ProspectsDocument19 pagesEffective Microbial Bioremediation Via The Multi-Omics Approach: An Overview of Trends, Problems and ProspectsUMYU Journal of Microbiology Research (UJMR)Pas encore d'évaluation
- Clot, Sánchez, Usán - Ferran, Elisabeth, Elsa - 2nlab - GSFP - P9F5 - Article Científic de Recerca FQDocument27 pagesClot, Sánchez, Usán - Ferran, Elisabeth, Elsa - 2nlab - GSFP - P9F5 - Article Científic de Recerca FQElisabethPas encore d'évaluation
- Antibiotics: Success and FailuresDocument43 pagesAntibiotics: Success and Failurestummalapalli venkateswara raoPas encore d'évaluation
- Ex3 Abo Forward Grouping Group7Document19 pagesEx3 Abo Forward Grouping Group7marielle rosePas encore d'évaluation
- Neural TubeDocument4 pagesNeural TubeRuth Danielle GasconPas encore d'évaluation
- Ribosom Kloroplas EMBJ 36 475Document12 pagesRibosom Kloroplas EMBJ 36 475NovyKedungWilutPas encore d'évaluation
- CHE EL3 Prelim Q Exam Feb 282024 Docx 1Document4 pagesCHE EL3 Prelim Q Exam Feb 282024 Docx 1MICHELLE MARIPOSAPas encore d'évaluation
- Introduction To Entomology: Definition-The Scientific Study of InsectsDocument25 pagesIntroduction To Entomology: Definition-The Scientific Study of Insectssadam hussainPas encore d'évaluation
- PROTISTDocument43 pagesPROTISTnurliyanaaminPas encore d'évaluation
- What Is DNA: FunctionDocument4 pagesWhat Is DNA: FunctionBartoszPas encore d'évaluation
- Hassan Et Al., 2024Document15 pagesHassan Et Al., 2024Herda CahyaningrumPas encore d'évaluation
- Antibacterial activities of the methanol extracts of Albizia adianthifolia, Alchornea laxiflora, Laportea ovalifolia and three other Cameroonian plants against multi-drug resistant Gram-negative bacteriaDocument6 pagesAntibacterial activities of the methanol extracts of Albizia adianthifolia, Alchornea laxiflora, Laportea ovalifolia and three other Cameroonian plants against multi-drug resistant Gram-negative bacteriaAriePas encore d'évaluation
- INTEGUMENTARY SYsTEMDocument42 pagesINTEGUMENTARY SYsTEMGabPas encore d'évaluation
- Module 1 Evolution of LifeDocument53 pagesModule 1 Evolution of LifeKeziah ChristinePas encore d'évaluation
- Nature at Work - The Ongoing Saga of Evolution - 400 PagsDocument400 pagesNature at Work - The Ongoing Saga of Evolution - 400 PagsNoti News WorldPas encore d'évaluation
- Digestive SystemDocument12 pagesDigestive SystemdelianamandhitaPas encore d'évaluation
- 7th Class Science-SEM2Document208 pages7th Class Science-SEM2Vishnu Vijay shankarPas encore d'évaluation
- Biochemistry and MetabolismDocument8 pagesBiochemistry and MetabolismPrincess Angie GonzalesPas encore d'évaluation