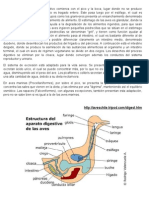
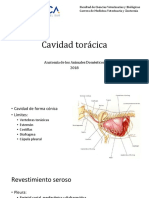
Vous aimerez peut-être aussi
- AbsorciÓn de Nutrientes en AvesDocument38 pagesAbsorciÓn de Nutrientes en AvesDiego Ruiz Ramos50% (8)
- Digestion y Absorcion de Lipidos en Monogastricos BlankaDocument45 pagesDigestion y Absorcion de Lipidos en Monogastricos BlankaJames Aguilar73% (15)
- Digestión de Carbohidratos en Monogástricos Pollo.Document8 pagesDigestión de Carbohidratos en Monogástricos Pollo.Mario Sergio Espinoza Aguilar100% (1)
- Ganglios LinfáticosDocument3 pagesGanglios LinfáticosJorge Andres Navarro0% (1)
- Sistema Respiratorio en Animales DomesticosDocument49 pagesSistema Respiratorio en Animales DomesticosLucianaMariñosCusquisibán77% (26)
- Anatomía del conejoDocument47 pagesAnatomía del conejoAndré MárquezPas encore d'évaluation
- Conceptualización y Análisis de Los Parámetros Reproductivos Del Equino CUARTO de MILLADocument26 pagesConceptualización y Análisis de Los Parámetros Reproductivos Del Equino CUARTO de MILLACristina HerreraPas encore d'évaluation
- Sistema - Digestivo Del Cerdo, Aves, Cuy y Cabras.Document24 pagesSistema - Digestivo Del Cerdo, Aves, Cuy y Cabras.DavidPas encore d'évaluation
- Teórico Hembra 2019-2Document96 pagesTeórico Hembra 2019-2Jennifer RodriguezPas encore d'évaluation
- INFORME COLECTA Semen EquinoDocument17 pagesINFORME COLECTA Semen Equinoandrea del pilarPas encore d'évaluation
- Anatomia Del ConejoDocument4 pagesAnatomia Del ConejoKaren Flores Ol67% (9)
- Aparato Digestivo de La OvejaDocument11 pagesAparato Digestivo de La Ovejas_rockosk8222Pas encore d'évaluation
- Anatomía Del Aparato Reproductor de La GallinaDocument3 pagesAnatomía Del Aparato Reproductor de La GallinaCristian Jasiel Barrios SuarezPas encore d'évaluation
- Alimentación y sistema digestivo de ovinos y caprinosDocument22 pagesAlimentación y sistema digestivo de ovinos y caprinosLoreanney Franco100% (1)
- Sem 2 Prod. de Porcinos-DDocument8 pagesSem 2 Prod. de Porcinos-DKram Ma Arm ArmasPas encore d'évaluation
- Desarrollo embrionario porcinoDocument5 pagesDesarrollo embrionario porcinoJhojan Victorio ChachiPas encore d'évaluation
- El Sistema Digestivo de La GallinaDocument6 pagesEl Sistema Digestivo de La GallinaslayporPas encore d'évaluation
- Anatomía y Fisiología Del Sistema Digestivo Del PorcinoDocument19 pagesAnatomía y Fisiología Del Sistema Digestivo Del PorcinoElwin Suarez Pájaro73% (11)
- Aparato Digestivo en Monogástrico y PoligástricoDocument35 pagesAparato Digestivo en Monogástrico y PoligástricoTalita Kumi82% (11)
- Metabolismo de Proteínas MonogástricoDocument26 pagesMetabolismo de Proteínas MonogástricoDesahily JuarezPas encore d'évaluation
- ConejosDocument42 pagesConejosJuand Neko VCruz50% (4)
- Necropsia en Conejos Autor JeanDocument9 pagesNecropsia en Conejos Autor JeanJean Carlos Quispe S0% (1)
- Cetosis Porcina 1Document13 pagesCetosis Porcina 1Eliel Ortiz100% (1)
- OvinoDocument6 pagesOvinoMaria Jose Hernandez0% (1)
- Anatomía y Fisiología Del GansoDocument6 pagesAnatomía y Fisiología Del GansoDarwin NavarroPas encore d'évaluation
- Requerimientos Nutricionales para CerdosDocument16 pagesRequerimientos Nutricionales para CerdosAlan AtoraPas encore d'évaluation
- Anatomia y Fisiologia Del CuyDocument5 pagesAnatomia y Fisiologia Del CuyGloria Stefani Diaz75% (16)
- Anatomia Del Sistemas Digestivo de Las Aves y PorcinosDocument8 pagesAnatomia Del Sistemas Digestivo de Las Aves y PorcinosLinda López Ruiz100% (1)
- Anatomía y Fisiología Del ConejoDocument8 pagesAnatomía y Fisiología Del ConejoOsmar Daniel Leguizamon0% (1)
- Digestion CamelidosDocument25 pagesDigestion CamelidosJonathan O. Mendoza Ch80% (5)
- Exposicion Grupal - Metabolismo de Los Carbohidratos en Monogastricos CaninosDocument9 pagesExposicion Grupal - Metabolismo de Los Carbohidratos en Monogastricos CaninosAndrea GarcesPas encore d'évaluation
- Procesos de Digestion de Las AvesDocument1 pageProcesos de Digestion de Las AvesAdrian José Perez SalazarPas encore d'évaluation
- Sistema Muscular de Las AvesDocument7 pagesSistema Muscular de Las AvesOmar Larios Maldonado88% (8)
- Anatomía y Fisiología Del Tracto Digestivo MonogástricosDocument66 pagesAnatomía y Fisiología Del Tracto Digestivo MonogástricosÓscar David Ramírez Mosquera100% (1)
- Criollo PillareñoDocument2 pagesCriollo PillareñoMonica GaleasPas encore d'évaluation
- Rol Del Agua en La Nutricion AnimalDocument17 pagesRol Del Agua en La Nutricion AnimalJhamil Lopez50% (2)
- Necropsia en AvesDocument5 pagesNecropsia en AvesSariel Pocota QuinterosPas encore d'évaluation
- Cecotrofia en Cuyes y ConejosDocument9 pagesCecotrofia en Cuyes y ConejosLuis Rengifo SaavedraPas encore d'évaluation
- Sistema Digestivo Del ConejoDocument23 pagesSistema Digestivo Del ConejoVeronica Matos Soriano87% (52)
- Aparato Digestivo Del CerdoDocument8 pagesAparato Digestivo Del CerdoJersonLimaymantaSalazarPas encore d'évaluation
- AparLocomoDocument21 pagesAparLocomoNubia Luna67% (3)
- Medidas Zoometricas en CerdosDocument16 pagesMedidas Zoometricas en CerdosJuanDAVIDS60% (5)
- Práctica Método de Dennis Stone SwansonDocument2 pagesPráctica Método de Dennis Stone SwansonElizabethPas encore d'évaluation
- Expo SemiologiaDocument78 pagesExpo SemiologiaSergio Andres Muñoz CollazosPas encore d'évaluation
- Carbohidratos en Nutricion EquinaDocument5 pagesCarbohidratos en Nutricion EquinaAlex D' RoorshackPas encore d'évaluation
- Sistema Digestivo Del Ovino-AnatomiaDocument5 pagesSistema Digestivo Del Ovino-AnatomiaMiguel Guillen100% (3)
- Anatomía cavidad torácica animalesDocument9 pagesAnatomía cavidad torácica animalesAnonymous 3yee0hOvG50% (2)
- La Espermatogenesis en El CerdoDocument36 pagesLa Espermatogenesis en El CerdoLuis Enrique Gonzales80% (5)
- Fisiología de La Puesta de La GallinaDocument12 pagesFisiología de La Puesta de La GallinaHenry Otoniel67% (3)
- Anatomia Del Conejo2Document53 pagesAnatomia Del Conejo2jaime_augusto_mvz@hotmail.com100% (3)
- Anatomía y Fisiología Del PavoDocument4 pagesAnatomía y Fisiología Del Pavoodlawsozepol100% (3)
- Constante Fisiologia AnimalDocument3 pagesConstante Fisiologia AnimalRaul Larios Curiel100% (2)
- Chabertia OvinaDocument5 pagesChabertia OvinaDuglaz Ramos DaySnot100% (1)
- El RUMIANTE Fisiología Digestiva y NutriciónDocument9 pagesEl RUMIANTE Fisiología Digestiva y NutriciónMicaela Colombo0% (1)
- Anatomía, Fisiología de La Digestión y Absorción de Carbohidratos PDFDocument4 pagesAnatomía, Fisiología de La Digestión y Absorción de Carbohidratos PDFEsauPas encore d'évaluation
- Digestion Absorcion CarbohidratosDocument4 pagesDigestion Absorcion CarbohidratosAndre Aylas100% (1)
- Fisiologia de Colon y CiegoDocument3 pagesFisiologia de Colon y CiegoFelipe Faura TrujilloPas encore d'évaluation
- 4 13-04, Resumen Ficha, ANATOMIA Y FISIOLOGIA DEL APARATO DIGESTIVO DEL CERDODocument7 pages4 13-04, Resumen Ficha, ANATOMIA Y FISIOLOGIA DEL APARATO DIGESTIVO DEL CERDOManuel Ulises VarelaPas encore d'évaluation
- Fisiología Digestiva en RumiantesDocument19 pagesFisiología Digestiva en RumiantesPablo Cesar Requena NeyraPas encore d'évaluation
- Modulo de Nutricion de Los Rumiantes PDFDocument162 pagesModulo de Nutricion de Los Rumiantes PDFfabioPas encore d'évaluation
- GRASP+V1 3 Interpretation+Ecuador+2017.en - EsDocument35 pagesGRASP+V1 3 Interpretation+Ecuador+2017.en - EsAle U. TitoañaPas encore d'évaluation
- Auditoría de Comercio Ético SMETADocument181 pagesAuditoría de Comercio Ético SMETAAle U. Titoaña100% (2)
- Guia Vida en El ExtranjeroDocument2 pagesGuia Vida en El ExtranjeroAle U. TitoañaPas encore d'évaluation
- PlanilladeinscripcionDocument1 pagePlanilladeinscripcionAle U. TitoañaPas encore d'évaluation
- Lista de Materiales PromedioDocument2 pagesLista de Materiales PromedioAle U. TitoañaPas encore d'évaluation
- Creencias LimitantesDocument1 pageCreencias LimitantesAle U. TitoañaPas encore d'évaluation
- La Importancia de Comer en FamiliaDocument1 pageLa Importancia de Comer en FamiliaAle U. TitoañaPas encore d'évaluation
- Proforma Emprosal PrigourmetDocument2 pagesProforma Emprosal PrigourmetAle U. TitoañaPas encore d'évaluation
- Ficha Tecnica Sal #5Document2 pagesFicha Tecnica Sal #5Ale U. TitoañaPas encore d'évaluation
- Regalos A PadresDocument1 pageRegalos A PadresAle U. TitoañaPas encore d'évaluation
- F01-F475-T Esparrago 2020 PDFDocument96 pagesF01-F475-T Esparrago 2020 PDFJUAN PABLOPas encore d'évaluation
- Esparrago U C 115Document1 pageEsparrago U C 115Ale U. TitoañaPas encore d'évaluation
- NORMATIVADocument4 pagesNORMATIVAAle U. TitoañaPas encore d'évaluation
- Preguntas Frecuentes de ECUAPASSDocument2 pagesPreguntas Frecuentes de ECUAPASSTodoComercioExteriorPas encore d'évaluation
- Convenio Pasantias General Espol-Astinave00010001Document8 pagesConvenio Pasantias General Espol-Astinave00010001Ale U. TitoañaPas encore d'évaluation
- Becas Israel 2018 en Español e InglésDocument2 pagesBecas Israel 2018 en Español e InglésAle U. TitoañaPas encore d'évaluation
- Memoria Curso Fertilización FoliarDocument145 pagesMemoria Curso Fertilización Foliarttonitta6956100% (2)
- Sistemas de Gestion de Calidad en El Sector AgroalimentarioDocument64 pagesSistemas de Gestion de Calidad en El Sector AgroalimentarioJose SerranoPas encore d'évaluation
- Manual InfostatDocument336 pagesManual InfostatlauseguraPas encore d'évaluation
- Temas Gym VirtualDocument9 pagesTemas Gym VirtualAle U. TitoañaPas encore d'évaluation
- Manejo Nutricional de Ponedoras en Fase de PosturaDocument3 pagesManejo Nutricional de Ponedoras en Fase de PosturaAle U. TitoañaPas encore d'évaluation
- Consejos para Sentirte Perfecta SDCDocument16 pagesConsejos para Sentirte Perfecta SDCAra Barrios67% (3)
- Nutrición Vegetal-Control de Lectura (Fósforo)Document2 pagesNutrición Vegetal-Control de Lectura (Fósforo)Ale U. TitoañaPas encore d'évaluation
- Poliploidía en peces: impactos en la acuiculturaDocument37 pagesPoliploidía en peces: impactos en la acuiculturaAle U. TitoañaPas encore d'évaluation
- Aspectos RealidadDocument3 pagesAspectos RealidadAle U. TitoañaPas encore d'évaluation
- Capítulo IDocument1 pageCapítulo IAle U. TitoañaPas encore d'évaluation
- Ppto Viv DS49-07-07-13 BNDocument36 pagesPpto Viv DS49-07-07-13 BNAle U. TitoañaPas encore d'évaluation
- HerbicidasDocument4 pagesHerbicidasAle U. TitoañaPas encore d'évaluation
- PedigriDocument1 pagePedigriAle U. TitoañaPas encore d'évaluation
- Analisis Sensorial LaboratorioDocument5 pagesAnalisis Sensorial LaboratorioStefy Ortiz BdvPas encore d'évaluation
- I Bimestre - 1° Grado - Activ 04Document6 pagesI Bimestre - 1° Grado - Activ 04Jhonny SalvadorPas encore d'évaluation
- Guion de ArteDocument3 pagesGuion de ArteJonaPas encore d'évaluation
- Guías alimentarias mundialesDocument9 pagesGuías alimentarias mundialesdaniel israel nava garcia100% (1)
- Caso Práctico LeyesDocument3 pagesCaso Práctico LeyesRebecca HdzPas encore d'évaluation
- Investicacion de La UvaDocument31 pagesInvesticacion de La UvaMarino Fernández SobradosPas encore d'évaluation
- Análisis CombinatorioDocument2 pagesAnálisis CombinatorioFranz Vasquez0% (1)
- Operaciones Unitarias de La Industria de Los AlimentosDocument36 pagesOperaciones Unitarias de La Industria de Los Alimentoscorpo8233% (3)
- Seguridad alimentaria en el Perú: Impacto de la pandemia y recomendacionesDocument17 pagesSeguridad alimentaria en el Perú: Impacto de la pandemia y recomendacionesRoger Salvatierra BreñaPas encore d'évaluation
- Estrategias de Dominio Neocolonial Detrás Del Concepto de Seguridad Alimentaria en VenezuelaDocument36 pagesEstrategias de Dominio Neocolonial Detrás Del Concepto de Seguridad Alimentaria en Venezuelacarlys lopezPas encore d'évaluation
- Diagrama de FlujoDocument4 pagesDiagrama de FlujoAndres Ivan Castilla UrianaPas encore d'évaluation
- MANUELITA FyH S.A.CDocument108 pagesMANUELITA FyH S.A.CMelberOmarCiezaTiradoPas encore d'évaluation
- Elaboracion Conservas de Pescado - Haccp, Poes y BPMDocument14 pagesElaboracion Conservas de Pescado - Haccp, Poes y BPMOmar Castillo100% (2)
- Estructuración Del Anteproyecto Y Proyecto de Investigación. Eje 2Document11 pagesEstructuración Del Anteproyecto Y Proyecto de Investigación. Eje 2MyriamSolarteAcostaPas encore d'évaluation
- Insecticidas BotánicosDocument35 pagesInsecticidas BotánicosDavidNuñezPas encore d'évaluation
- NJKKDocument12 pagesNJKKAstridPas encore d'évaluation
- Encuesta Miel de AbejaDocument5 pagesEncuesta Miel de AbejaGradiz Murillo100% (2)
- 84 Recetas de Comida Express - OrzolaDocument98 pages84 Recetas de Comida Express - Orzolayorlen100% (1)
- Diapositivas de RestauranteDocument16 pagesDiapositivas de RestauranteErnestoNuñezPas encore d'évaluation
- Jugo de Naranja 04-05-2009Document3 pagesJugo de Naranja 04-05-2009William Zavaleta Garcia100% (1)
- Alcachofa Parte AgricolaDocument43 pagesAlcachofa Parte AgricolaChristian TuniPas encore d'évaluation
- Guia 1 Matematicas 1 Uan 2019 1Document6 pagesGuia 1 Matematicas 1 Uan 2019 1Dylan RamirezPas encore d'évaluation
- El Magnesio y Sus Propiedades CurativasDocument62 pagesEl Magnesio y Sus Propiedades Curativasjose guevaraPas encore d'évaluation
- 20 Canciones ImprimirDocument5 pages20 Canciones ImprimirMartina TrujilloPas encore d'évaluation
- Receta Estandar PDFDocument4 pagesReceta Estandar PDFgaboreyes33% (3)
- DIAPOS DE Camu Camu 3Document18 pagesDIAPOS DE Camu Camu 3Carlos GasteloPas encore d'évaluation
- Guía nutricional para pacientes oncológicosDocument12 pagesGuía nutricional para pacientes oncológicosjohan sebastian valencia marinPas encore d'évaluation
- Proy. Cientifico Insectisida A Base de Ajo para Los Huertos Escolares Leydi Prima1Document46 pagesProy. Cientifico Insectisida A Base de Ajo para Los Huertos Escolares Leydi Prima1Humberto Montoya100% (2)
- InstrumentosDocument4 pagesInstrumentosNiikii Jara VelascoPas encore d'évaluation