Vous aimerez peut-être aussi
- Presented By: Jet Daniel Andrei Arellano Darwin Reigh Calub Aeryl Dierson MagahisDocument75 pagesPresented By: Jet Daniel Andrei Arellano Darwin Reigh Calub Aeryl Dierson MagahisDaniel Arellano85% (13)
- AcidoDocument6 pagesAcidoDark WolfPas encore d'évaluation
- Cells: Fferential Regulation of Circrna, Mirna, and Fferentiation of Human Mesenchymal Stromal CellsDocument24 pagesCells: Fferential Regulation of Circrna, Mirna, and Fferentiation of Human Mesenchymal Stromal CellsBreeze ChloePas encore d'évaluation
- Caco Cell LineDocument9 pagesCaco Cell Linedonaldozc07Pas encore d'évaluation
- Jahr Et Al., 2005Document6 pagesJahr Et Al., 2005Angelescu LauraPas encore d'évaluation
- The Spatial Transcriptomic Landscape of The Healing Mouse Intestine Following DamageDocument16 pagesThe Spatial Transcriptomic Landscape of The Healing Mouse Intestine Following Damageyjbu0125Pas encore d'évaluation
- Phosphatidylinositol 3 - Kinase Signalling Supports Cell Height in Established Epithelial MonolayersDocument11 pagesPhosphatidylinositol 3 - Kinase Signalling Supports Cell Height in Established Epithelial MonolayersMohammed Elias AlamPas encore d'évaluation
- Publikacja 3Document13 pagesPublikacja 3Przemysław ZakrzewskiPas encore d'évaluation
- MainDocument8 pagesMainAxel MendozaPas encore d'évaluation
- Expression of A Gene Encoding Trypanosoma Congolense Putative Abc1 Family Protein Is Developmentally RegulatedDocument0 pageExpression of A Gene Encoding Trypanosoma Congolense Putative Abc1 Family Protein Is Developmentally RegulatedMaísa FonsecaPas encore d'évaluation
- Mitochondrial Energy Metabolism in Baby Hamster Kidney (BHK-21/C13) Cells Treated With Karnozin EXTRA®Document7 pagesMitochondrial Energy Metabolism in Baby Hamster Kidney (BHK-21/C13) Cells Treated With Karnozin EXTRA®Garbuz ElenaPas encore d'évaluation
- tmpD776 TMPDocument6 pagestmpD776 TMPFrontiersPas encore d'évaluation
- Art 1Document18 pagesArt 1Luis Hurtado PardoPas encore d'évaluation
- 1 s2.0 S0014299903027250 MainDocument8 pages1 s2.0 S0014299903027250 MainZulfiani SyachbaniahPas encore d'évaluation
- Tissue Engineering and Regeneration in Dentistry: Current StrategiesD'EverandTissue Engineering and Regeneration in Dentistry: Current StrategiesRachel J. WaddingtonPas encore d'évaluation
- Engleska Veryija RadaDocument6 pagesEngleska Veryija RadaNevena SaulicPas encore d'évaluation
- Amorphous, Smart, and Bioinspired Polyphosphate Nano/Microparticles: A Biomaterial For Regeneration and Repair of Osteo-Articular Impairments In-SituDocument19 pagesAmorphous, Smart, and Bioinspired Polyphosphate Nano/Microparticles: A Biomaterial For Regeneration and Repair of Osteo-Articular Impairments In-SituMiftah NajihPas encore d'évaluation
- 1-S2.0-S0929664621001923-Main (2) Pulp LTDocument8 pages1-S2.0-S0929664621001923-Main (2) Pulp LTKena BhalaniPas encore d'évaluation
- mRNA and Long Non-Coding RNA Expression Profiling of Human Periodontal Ligament Cells Under Tension LoadingDocument10 pagesmRNA and Long Non-Coding RNA Expression Profiling of Human Periodontal Ligament Cells Under Tension LoadingJuliana ÁlvarezPas encore d'évaluation
- 1 s2.0 S1465324917300051 MainDocument15 pages1 s2.0 S1465324917300051 MainPando RaitaPas encore d'évaluation
- Immortalized Liver Endothel CCmodelDocument20 pagesImmortalized Liver Endothel CCmodelAndreiPas encore d'évaluation
- Bcri2012 672705Document14 pagesBcri2012 672705Sergeat18BPas encore d'évaluation
- NIH Public Access: Author ManuscriptDocument8 pagesNIH Public Access: Author ManuscriptAhmad AbdelFattah HafezPas encore d'évaluation
- Kuze 1997Document8 pagesKuze 1997Sarly FebrianaPas encore d'évaluation
- Ijmm 25 4 593 PDFDocument8 pagesIjmm 25 4 593 PDFNickPas encore d'évaluation
- Tussilagone Promotes Osteoclast Apoptosis and Prevents Estrogen Deficiency-Induced Osteoporosis in MiceDocument7 pagesTussilagone Promotes Osteoclast Apoptosis and Prevents Estrogen Deficiency-Induced Osteoporosis in MiceMAGALI GONZÁLEZ RODRÍGUEZPas encore d'évaluation
- BMC Biotechnology: Dynamic in Vivo Imaging and Cell Tracking Using A Histone Fluorescent Protein Fusion in MiceDocument14 pagesBMC Biotechnology: Dynamic in Vivo Imaging and Cell Tracking Using A Histone Fluorescent Protein Fusion in Micefather45Pas encore d'évaluation
- Integrative CUT&Tag-RNA-Seq Analysis of Histone Variant macroH2A1-dependent Orchestration of Human Induced Pluripotent Stem Cell ReprogrammingDocument16 pagesIntegrative CUT&Tag-RNA-Seq Analysis of Histone Variant macroH2A1-dependent Orchestration of Human Induced Pluripotent Stem Cell Reprogrammingniccolo.liorni1Pas encore d'évaluation
- The Eutt Gene of Salmonella Enterica Encodes An Oxygen-Labile, Metal-Containing ATP:Corrinoid Adenosyltransferase EnzymeDocument9 pagesThe Eutt Gene of Salmonella Enterica Encodes An Oxygen-Labile, Metal-Containing ATP:Corrinoid Adenosyltransferase EnzymeEdwardPas encore d'évaluation
- 1953 FullDocument10 pages1953 FullboboyoyotPas encore d'évaluation
- Inter-Cellular Nanovesicle Mediated MicroRNA TransferDocument19 pagesInter-Cellular Nanovesicle Mediated MicroRNA Transferanon_579968442Pas encore d'évaluation
- CytometryDocument11 pagesCytometrymitkazPas encore d'évaluation
- Visualization and Functional Analysis of A Maxi-K Channel (Mslo) Fused To Green Fluorescent Protein (GFP)Document12 pagesVisualization and Functional Analysis of A Maxi-K Channel (Mslo) Fused To Green Fluorescent Protein (GFP)sadfdsfPas encore d'évaluation
- Tunable Crosslinked Cell Derived Extracellular Matrix Guides Cell FateDocument12 pagesTunable Crosslinked Cell Derived Extracellular Matrix Guides Cell FateMuhammad SuhaeriPas encore d'évaluation
- Kawasaki 1997Document6 pagesKawasaki 1997AmatystPas encore d'évaluation
- Isolation and Characterisation of Mesenchymal Stem Cells From Adult Mouse Bone MarrowDocument12 pagesIsolation and Characterisation of Mesenchymal Stem Cells From Adult Mouse Bone MarrowRamasamy19Pas encore d'évaluation
- 39 809Document7 pages39 809windyndaPas encore d'évaluation
- Electrical Characters On Surface of Apoptotic Cell in Maize Roots Induced by CytotoxinsDocument10 pagesElectrical Characters On Surface of Apoptotic Cell in Maize Roots Induced by CytotoxinsUmiatin RamdhaniPas encore d'évaluation
- Induced Monocytes-Derived Hscs (Cd34+) With Lps Accelerated Homing Rat Bone Marrow-Mesenchymal Stem Cell (BM-MSCS, Cd105) in Injured PancreasDocument12 pagesInduced Monocytes-Derived Hscs (Cd34+) With Lps Accelerated Homing Rat Bone Marrow-Mesenchymal Stem Cell (BM-MSCS, Cd105) in Injured PancreasAzmi FarhadiPas encore d'évaluation
- Bone Marrow Stromal Cells Produce Thrombopoietin and Stimulate Megakaryocyte Growth and Maturation But Suppress Proplatelet FormationDocument8 pagesBone Marrow Stromal Cells Produce Thrombopoietin and Stimulate Megakaryocyte Growth and Maturation But Suppress Proplatelet FormationLeonardo SalesPas encore d'évaluation
- (Nanoparticles-Based Drug Delivery Systems (NDDSS) or Biomimetic Nanoparticles) Tumor Exosome-Based Nanoparticles For ChemotherapyDocument16 pages(Nanoparticles-Based Drug Delivery Systems (NDDSS) or Biomimetic Nanoparticles) Tumor Exosome-Based Nanoparticles For ChemotherapysylodhiPas encore d'évaluation
- Stirred Suspension Bioreactor Culture of Porcine Induced Pluripotent Stem CellsDocument12 pagesStirred Suspension Bioreactor Culture of Porcine Induced Pluripotent Stem CellsArianna CastroPas encore d'évaluation
- Comparing Two Intestinal Porcine Epithelial Cell Lines (Ipecs) : Morphological Differentiation, Function and MetabolismDocument20 pagesComparing Two Intestinal Porcine Epithelial Cell Lines (Ipecs) : Morphological Differentiation, Function and MetabolismCristina BulgaruPas encore d'évaluation
- Expression of Bone Morphogenetic Proteins in Giant Cell Tumor of BoneDocument7 pagesExpression of Bone Morphogenetic Proteins in Giant Cell Tumor of BoneAyu SugiartyPas encore d'évaluation
- Metabolic Fingerprints of Human Primary Endothelial and Fibroblast CellsDocument12 pagesMetabolic Fingerprints of Human Primary Endothelial and Fibroblast CellsDavid Galiano LatorrePas encore d'évaluation
- Endotelial Pericites - Cui2006Document10 pagesEndotelial Pericites - Cui2006Lilo FrancoisPas encore d'évaluation
- In Vitro Zonation and Toxicity in A Hepatocyte Bioreactor: Toxicological SciencesDocument10 pagesIn Vitro Zonation and Toxicity in A Hepatocyte Bioreactor: Toxicological SciencesRamboPas encore d'évaluation
- Braga C Cytotherapy 2022 Proteomics of MSC EV Normoxia HypoxiaDocument14 pagesBraga C Cytotherapy 2022 Proteomics of MSC EV Normoxia HypoxiaAlmira AlodiaPas encore d'évaluation
- Rank - BPsDocument8 pagesRank - BPsIsteicy CortezPas encore d'évaluation
- AKT/Protein Kinase B Regulation of BCL Family Members During Oxysterol-Induced ApoptosisDocument8 pagesAKT/Protein Kinase B Regulation of BCL Family Members During Oxysterol-Induced ApoptosisNareshPas encore d'évaluation
- FOXK1 Plays An Oncogenic Role in The Deve 2017 Biochemical and Biophysical RDocument7 pagesFOXK1 Plays An Oncogenic Role in The Deve 2017 Biochemical and Biophysical RFlorin AchimPas encore d'évaluation
- Life Sciences: A B A A A ADocument10 pagesLife Sciences: A B A A A AsovalaxPas encore d'évaluation
- Poster-Presentaion-Abstracts ASPIRE2017Document241 pagesPoster-Presentaion-Abstracts ASPIRE2017HAVIZ YUADPas encore d'évaluation
- Unveiling Novel Genes Upregulated by Both Rhbmp2 and Rhbmp7 During Early Osteoblastic Transdifferentiation of C2C12 CellsDocument15 pagesUnveiling Novel Genes Upregulated by Both Rhbmp2 and Rhbmp7 During Early Osteoblastic Transdifferentiation of C2C12 Cellsvanessa_werbickyPas encore d'évaluation
- Articulo2 1Document36 pagesArticulo2 1DanielaSofiaRubianoBlancoPas encore d'évaluation
- Dex3 and Survivin-2B: Two Novel Splice Variants of The ApoptosisDocument6 pagesDex3 and Survivin-2B: Two Novel Splice Variants of The ApoptosisNona NonicaaPas encore d'évaluation
- Hybrid Embryonic Stem Cell-Derived Tetraploid Mice Show Apparently Normal Morphological, Physiological, and Neurological CharacteristicsDocument8 pagesHybrid Embryonic Stem Cell-Derived Tetraploid Mice Show Apparently Normal Morphological, Physiological, and Neurological CharacteristicsGmewop30mPas encore d'évaluation
- Extracellular Vesicles (Ber)Document13 pagesExtracellular Vesicles (Ber)Farhan Royan PermanahadiPas encore d'évaluation
- LNC NTF3 5 Promotes Osteogenic Differentiation of Maxillary Sinus Membrane Stem Cells Via Sponging Mir 93 3pDocument12 pagesLNC NTF3 5 Promotes Osteogenic Differentiation of Maxillary Sinus Membrane Stem Cells Via Sponging Mir 93 3pBagis Emre GulPas encore d'évaluation
- 9 Flow Cytometric Strategies To Study CNS DevelopmentDocument32 pages9 Flow Cytometric Strategies To Study CNS DevelopmenttaroPas encore d'évaluation
- Carbon Nanotubes As Intracellular Protein Transporters: Generality and Biological FunctionalityDocument6 pagesCarbon Nanotubes As Intracellular Protein Transporters: Generality and Biological Functionality227986Pas encore d'évaluation
- Sexuality and Spinal Cord InjuryDocument10 pagesSexuality and Spinal Cord InjurySebastián GallegosPas encore d'évaluation
- Direct and Indirect Pathways of Basal Ganglia A Critical ReppraisalDocument9 pagesDirect and Indirect Pathways of Basal Ganglia A Critical ReppraisalSebastián Gallegos0% (1)
- Neuromuscular Manifestations of Critical Illness: Invited ReviewDocument24 pagesNeuromuscular Manifestations of Critical Illness: Invited ReviewSebastián GallegosPas encore d'évaluation
- Cognitive Impairment and Dementia in Parkinson's Disease - Clinical Features, Diagnosis, and ManagementDocument15 pagesCognitive Impairment and Dementia in Parkinson's Disease - Clinical Features, Diagnosis, and ManagementSebastián GallegosPas encore d'évaluation
- Basal Ganglia Circuits As Targets For Neuromodulation in Parkinson DiseaseDocument7 pagesBasal Ganglia Circuits As Targets For Neuromodulation in Parkinson DiseaseSebastián GallegosPas encore d'évaluation
- Brain: Altering Spinal Cord Excitability Enables Voluntary Movements After Chronic Complete Paralysis in HumansDocument16 pagesBrain: Altering Spinal Cord Excitability Enables Voluntary Movements After Chronic Complete Paralysis in HumansSebastián GallegosPas encore d'évaluation
- Osteogenesis Imperfecta: Epidemiology and Pathophysiology: Elizabeth Martin, MHS, and Jay R. Shapiro, MDDocument7 pagesOsteogenesis Imperfecta: Epidemiology and Pathophysiology: Elizabeth Martin, MHS, and Jay R. Shapiro, MDSebastián GallegosPas encore d'évaluation
- Katz 2002 VisualPlasticity HW 4Document9 pagesKatz 2002 VisualPlasticity HW 4Sebastián GallegosPas encore d'évaluation
- The Structure of Multi-Neuron Firing Patterns in Primate RetinaDocument13 pagesThe Structure of Multi-Neuron Firing Patterns in Primate RetinaSebastián GallegosPas encore d'évaluation
- Annurev - Castillo PDocument27 pagesAnnurev - Castillo PSebastián GallegosPas encore d'évaluation
- Mount Castle 1957Document27 pagesMount Castle 1957Sebastián GallegosPas encore d'évaluation
- Thomson 2007 Functional Maps of Neocortical CircuitryDocument24 pagesThomson 2007 Functional Maps of Neocortical CircuitrySebastián GallegosPas encore d'évaluation
- Neuroscience PDFDocument4 pagesNeuroscience PDFSebastián GallegosPas encore d'évaluation
- 599 FullDocument4 pages599 FullSebastián GallegosPas encore d'évaluation
- Experiments With Planaria PDFDocument40 pagesExperiments With Planaria PDFFrancisco Girbal Eiras0% (1)
- Functional Matrix HypothesisDocument16 pagesFunctional Matrix HypothesisUddyot ChaudharyPas encore d'évaluation
- Physiology of Labor PDFDocument68 pagesPhysiology of Labor PDFAJ MendozaPas encore d'évaluation
- C. R. Green (Auth.), Peter A. v. Anderson (Eds.) - Evolution of The First Nervous Systems-Springer US (1989)Document427 pagesC. R. Green (Auth.), Peter A. v. Anderson (Eds.) - Evolution of The First Nervous Systems-Springer US (1989)Vlad TomaPas encore d'évaluation
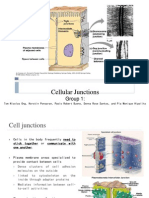
- Cellular JunctionsDocument25 pagesCellular JunctionsDiana Grace Pile0% (1)
- Cell Junction: 3 Types Intercellular Junctions in AnimalsDocument7 pagesCell Junction: 3 Types Intercellular Junctions in AnimalsNada hasanPas encore d'évaluation
- Type of Cell Junctions 1Document12 pagesType of Cell Junctions 1shahrul mukminPas encore d'évaluation
- Functional Matrix Revisited 2 PDFDocument6 pagesFunctional Matrix Revisited 2 PDFPranshu MathurPas encore d'évaluation
- Patel 2014Document7 pagesPatel 2014Javiera Munizaga MuñozPas encore d'évaluation
- Biosystems: Javier Cervera, Michael Levin, Salvador MafeDocument6 pagesBiosystems: Javier Cervera, Michael Levin, Salvador MafeAngelica Maria Torregroza EspinosaPas encore d'évaluation
- The Inferior Olivary Nucleus of The Rat A Light and Electron Microscopic StudyDocument31 pagesThe Inferior Olivary Nucleus of The Rat A Light and Electron Microscopic StudyRafael GutierrezPas encore d'évaluation
- Komunikasi Antar Sel: Irma Yuniar Wardhani, M.PDDocument12 pagesKomunikasi Antar Sel: Irma Yuniar Wardhani, M.PDIrma YuniarPas encore d'évaluation
- Basis of Chloride Transport in Ciliary EpitheliumDocument13 pagesBasis of Chloride Transport in Ciliary EpitheliumShashi kant ChaudharyPas encore d'évaluation
- 2009-High Resolution Map of Caenorhabditis Elegans Gap Junction ProteinsDocument15 pages2009-High Resolution Map of Caenorhabditis Elegans Gap Junction ProteinsyicinenPas encore d'évaluation
- Functional Matrix HypothesisDocument14 pagesFunctional Matrix Hypothesispriyab710Pas encore d'évaluation
- The Physiological Foundation of Yoga Chakra ExpressionDocument19 pagesThe Physiological Foundation of Yoga Chakra ExpressionTrev NeoPas encore d'évaluation
- Cell Junctions, Cell-Cell AdhesionsDocument36 pagesCell Junctions, Cell-Cell AdhesionsRizky Nur IndahPas encore d'évaluation
- Moss Functional Matrix RevisitedDocument23 pagesMoss Functional Matrix RevisitedPriyank RaiPas encore d'évaluation
- The Retinal Muller Cell PDFDocument293 pagesThe Retinal Muller Cell PDFkemalasari8818Pas encore d'évaluation
- Cellular Small Talk.Document8 pagesCellular Small Talk.Carlos VelascoPas encore d'évaluation