Vous aimerez peut-être aussi
- Pulse Crop Effects On Gut Microbial Populations, Intestinal Function, and Adiposity in A Mouse Model of Diet-Induced Obesity John N. McginleyDocument40 pagesPulse Crop Effects On Gut Microbial Populations, Intestinal Function, and Adiposity in A Mouse Model of Diet-Induced Obesity John N. Mcginleyzora.mendez637100% (8)
- Nutrients 05 01417 With CoverDocument20 pagesNutrients 05 01417 With Covervelkov.vladiPas encore d'évaluation
- Impact of Banana Puree On Viability of P PDFDocument8 pagesImpact of Banana Puree On Viability of P PDFAndreea FlorentinaPas encore d'évaluation
- Nutrients 13 03965 v2Document11 pagesNutrients 13 03965 v2VaniPas encore d'évaluation
- In Vitro SourdoughDocument11 pagesIn Vitro SourdoughMaryam JameelahPas encore d'évaluation
- BiavatiB Probiotics and PrebioticsDocument75 pagesBiavatiB Probiotics and PrebioticsRashmin ShahPas encore d'évaluation
- Prebiotic Effects of Partially Hydrolyzed Guar Gum On The Composition and Function of The Human Microbiota-Results From The PAGODA TrialDocument15 pagesPrebiotic Effects of Partially Hydrolyzed Guar Gum On The Composition and Function of The Human Microbiota-Results From The PAGODA TrialLevente BalázsPas encore d'évaluation
- Fermented Products With Probiotic Qualities - Casei & Bifidobakterije PDFDocument6 pagesFermented Products With Probiotic Qualities - Casei & Bifidobakterije PDFmilu1312Pas encore d'évaluation
- Aquaculture: Jiqiu Li, Beiping Tan, Kangsen MaiDocument6 pagesAquaculture: Jiqiu Li, Beiping Tan, Kangsen MaiOana Mariana IonetePas encore d'évaluation
- DR Neerja Hajela PPT Probiotics Prebiotics ILSI FinalDocument29 pagesDR Neerja Hajela PPT Probiotics Prebiotics ILSI Finaljbabu123Pas encore d'évaluation
- Jurnal Healthy: Disusun Dalam Rangka Memenuhi Tugas Mata Kuliah Bahasa Inggris Dosen Pembimbing: Lia Febriani, M.PDDocument6 pagesJurnal Healthy: Disusun Dalam Rangka Memenuhi Tugas Mata Kuliah Bahasa Inggris Dosen Pembimbing: Lia Febriani, M.PDandrye pelitaPas encore d'évaluation
- Effects of Dietary Yogurt On The Healthy Human Gastrointestinal (GI) MicrobiomeDocument16 pagesEffects of Dietary Yogurt On The Healthy Human Gastrointestinal (GI) Microbiomejasson ortega brionesPas encore d'évaluation
- FOS LectureDocument15 pagesFOS LectureernestoPas encore d'évaluation
- Five Tips To Help Cultivate A Healthy MicrobiomeDocument9 pagesFive Tips To Help Cultivate A Healthy MicrobiomeCristina Дима100% (1)
- Health Benefits of Prebiotics and Probiotics: Specialty "Food "Document18 pagesHealth Benefits of Prebiotics and Probiotics: Specialty "Food "Alexandra Ioan100% (1)
- Significance of Fermented Food in Nutrition and Food ScienceDocument14 pagesSignificance of Fermented Food in Nutrition and Food Scienceoryzae sativaPas encore d'évaluation
- Beneficial Effect of Intestinal Fermentation ofDocument22 pagesBeneficial Effect of Intestinal Fermentation ofLaszloPas encore d'évaluation
- Dietary Fibre Functionality in Food and Nutraceuticals: From Plant to GutD'EverandDietary Fibre Functionality in Food and Nutraceuticals: From Plant to GutFarah HosseinianPas encore d'évaluation
- Efectos en NiñosDocument8 pagesEfectos en NiñosSergio Benavides OcampoPas encore d'évaluation
- Review 2 ProbioticsDocument4 pagesReview 2 ProbioticsJose Oscar Mantilla RivasPas encore d'évaluation
- Nutrition JournalDocument7 pagesNutrition JournalLateecka R KulkarniPas encore d'évaluation
- Concise Review: Importance of Probiotics Yogurt For Human Health ImprovementDocument6 pagesConcise Review: Importance of Probiotics Yogurt For Human Health ImprovementIOSRjournalPas encore d'évaluation
- Fermented Foods and Beverages in Human Diet and Their Influence On Gut Microbiota and HealthDocument13 pagesFermented Foods and Beverages in Human Diet and Their Influence On Gut Microbiota and HealthDamar MurtiPas encore d'évaluation
- Assignment Pre and ProDocument14 pagesAssignment Pre and ProLatifah AhmadPas encore d'évaluation
- 1 s2.0 S1756464619302737 MainDocument12 pages1 s2.0 S1756464619302737 MainZhenquan JiaPas encore d'évaluation
- Dietary Interventions in Gastrointestinal Diseases: Foods, Nutrients, and Dietary SupplementsD'EverandDietary Interventions in Gastrointestinal Diseases: Foods, Nutrients, and Dietary SupplementsPas encore d'évaluation
- Effect of Probiotic and Prebiotic As Antibiotic Growth Promoter Substitutions On Productive and Carcass Traits of Broiler ChicksDocument5 pagesEffect of Probiotic and Prebiotic As Antibiotic Growth Promoter Substitutions On Productive and Carcass Traits of Broiler ChicksSureshCoolPas encore d'évaluation
- Thesis On Functional FoodsDocument6 pagesThesis On Functional FoodsPaySomeoneToWriteAPaperForMeCanada100% (2)
- Nutritional Value of Spirulina and Its U PDFDocument21 pagesNutritional Value of Spirulina and Its U PDFNel CasupangPas encore d'évaluation
- Patricia ProjectDocument22 pagesPatricia ProjectAkamshu GabrielPas encore d'évaluation
- FNS20110500005 33779711Document6 pagesFNS20110500005 33779711fannisaputriPas encore d'évaluation
- Aj 1Document11 pagesAj 1Trần Tấn TàiPas encore d'évaluation
- 1 s2.0 S0022030210004868 Main PDFDocument10 pages1 s2.0 S0022030210004868 Main PDFAliffiaPas encore d'évaluation
- Regular Egg Consumption at Breakfast by Japanese Woman University Students Improves Daily Nutrient Intakes: Open-Labeled ObservationsDocument7 pagesRegular Egg Consumption at Breakfast by Japanese Woman University Students Improves Daily Nutrient Intakes: Open-Labeled ObservationsLeena MuniandyPas encore d'évaluation
- Author's Accepted Manuscript: Food BioscienceDocument39 pagesAuthor's Accepted Manuscript: Food BiosciencerenPas encore d'évaluation
- Nutrients 12 02474Document15 pagesNutrients 12 02474Robert StryjakPas encore d'évaluation
- Diet-Microbe Interactions in the Gut: Effects on Human Health and DiseaseD'EverandDiet-Microbe Interactions in the Gut: Effects on Human Health and DiseaseKieran TuohyÉvaluation : 4.5 sur 5 étoiles4.5/5 (2)
- Apresentação de ArtigoDocument8 pagesApresentação de ArtigoJéssica BaumwollePas encore d'évaluation
- Probiotics - Friendly Bacteria With A Host of Benefits: HistoryDocument4 pagesProbiotics - Friendly Bacteria With A Host of Benefits: Historynikunj_patel_81Pas encore d'évaluation
- Intestinal Colonisation, Microbiota and Future Probiotics?: Original ArticleDocument5 pagesIntestinal Colonisation, Microbiota and Future Probiotics?: Original ArticleSherif Abou BakrPas encore d'évaluation
- 1 s2.0 S0271531712000176 MainDocument8 pages1 s2.0 S0271531712000176 MainEdwin RizoPas encore d'évaluation
- 6 IFRJ 19 (04) 2012 ShahDocument14 pages6 IFRJ 19 (04) 2012 ShahmarinaPas encore d'évaluation
- Prebioticsand Inflammatorybowel Disease: Heather E. Rasmussen,, Bruce R. HamakerDocument13 pagesPrebioticsand Inflammatorybowel Disease: Heather E. Rasmussen,, Bruce R. HamakerDiogo SouzaPas encore d'évaluation
- Probiotics Lesson PlanDocument7 pagesProbiotics Lesson Planapi-272776993Pas encore d'évaluation
- Prebiotic Effects of Inulin and Acacia GumDocument9 pagesPrebiotic Effects of Inulin and Acacia GumЃоре НаковPas encore d'évaluation
- IFRJ 2017, Sri Winarti, Colonic MicrobiotaDocument8 pagesIFRJ 2017, Sri Winarti, Colonic MicrobiotaSri WinartiPas encore d'évaluation
- Polysaccharides - Bowel Health and Gut MicrobiotaDocument14 pagesPolysaccharides - Bowel Health and Gut MicrobiotaLaszlo100% (1)
- Synbiotics: A Technological Approach in Food ApplicationsDocument14 pagesSynbiotics: A Technological Approach in Food ApplicationsJuan Fernando Cedillo RodríguezPas encore d'évaluation
- Acosta-Coello 2021Document12 pagesAcosta-Coello 2021Mariana SorianoPas encore d'évaluation
- MNM211544 NVDocument15 pagesMNM211544 NVBOUABSAPas encore d'évaluation
- 8 - Kefir ImproveDocument11 pages8 - Kefir ImproveCecep SanusiPas encore d'évaluation
- Prebiotik ProbiotikDocument8 pagesPrebiotik ProbiotikEka PutriPas encore d'évaluation
- 1 s2.0 S0308814623016242 MainDocument11 pages1 s2.0 S0308814623016242 MainJ CPas encore d'évaluation
- JFNR 4 3 7Document8 pagesJFNR 4 3 7AlbertochoPas encore d'évaluation
- Roquette - PrebioticDocument6 pagesRoquette - PrebiotichymasarathiPas encore d'évaluation
- Os Efeitos Do Comportamento Murino de Lactobacillus Helveticuson Dependem Da Dieta e GenótipoDocument10 pagesOs Efeitos Do Comportamento Murino de Lactobacillus Helveticuson Dependem Da Dieta e Genótipoj238111Pas encore d'évaluation
- The Benefits of Gene TherapyDocument3 pagesThe Benefits of Gene TherapyLevi AckermanPas encore d'évaluation
- Functional Foods.: Separating Fact From FictionDocument43 pagesFunctional Foods.: Separating Fact From Fictionmedical studiesPas encore d'évaluation
- Chemical Composition of Some Selected Fruit PeelsDocument10 pagesChemical Composition of Some Selected Fruit PeelsDaniel David ArangurenPas encore d'évaluation
- Crutzen2002 PDFDocument1 pageCrutzen2002 PDFDaniel David ArangurenPas encore d'évaluation
- Vocabulary - To Kill A MockingbirdDocument2 pagesVocabulary - To Kill A MockingbirdDaniel David ArangurenPas encore d'évaluation
- Batch Drying of Banana: Modelling and ExperimentsDocument6 pagesBatch Drying of Banana: Modelling and ExperimentsDaniel David ArangurenPas encore d'évaluation
- Who Donate BloodDocument4 pagesWho Donate BloodmanojPas encore d'évaluation
- Hiv and Aids - ReportDocument8 pagesHiv and Aids - Reportchezyl cadinongPas encore d'évaluation
- Small For Gestational Age (SGA)Document3 pagesSmall For Gestational Age (SGA)Adiel CalsaPas encore d'évaluation
- Vesiculobullous LesionDocument31 pagesVesiculobullous Lesionsara ibrahimPas encore d'évaluation
- Brochure in Health Ed PDFDocument3 pagesBrochure in Health Ed PDFHannalene DolorPas encore d'évaluation
- Senior Pet Nutrition and ManagementDocument17 pagesSenior Pet Nutrition and ManagementMayteMGPas encore d'évaluation
- What Causes EpilepsyDocument2 pagesWhat Causes EpilepsyJames CollinsPas encore d'évaluation
- Patient - Carpal Tunnel ReleaseDocument2 pagesPatient - Carpal Tunnel Releaseapi-277825138Pas encore d'évaluation
- Paul Zane PilzerDocument5 pagesPaul Zane PilzerJake Petrykowski50% (2)
- HypocalcemiaDocument23 pagesHypocalcemiarayrrn00Pas encore d'évaluation
- Skew Deviation RevisitedDocument24 pagesSkew Deviation RevisitedPriscila Verduzco MartínezPas encore d'évaluation
- Reading and WritingDocument9 pagesReading and WritingDECEDERIO CANDOLEPas encore d'évaluation
- Homocysteine and Markers of Coagulation and Endothelial Cell ActivationDocument7 pagesHomocysteine and Markers of Coagulation and Endothelial Cell ActivationMaya RustamPas encore d'évaluation
- Cholelithiasis: Group MembersDocument12 pagesCholelithiasis: Group MembersShiela GutierrezPas encore d'évaluation
- Stress (Biology) - Wikipedia, The Free Encyclopedia PDFDocument8 pagesStress (Biology) - Wikipedia, The Free Encyclopedia PDFBruno JeronimoPas encore d'évaluation
- The Role of EEG in Epilepsy - A Critical ReviewDocument12 pagesThe Role of EEG in Epilepsy - A Critical ReviewNicki Pinzon NariñoPas encore d'évaluation
- Cyto 18 Feb 2021Document6 pagesCyto 18 Feb 2021satria92Pas encore d'évaluation
- SB 5293 3 1Document2 pagesSB 5293 3 1api-659209201Pas encore d'évaluation
- Parathyroid Glands: Serum PTH Levels Are Inappropriately Elevated For The LevelDocument4 pagesParathyroid Glands: Serum PTH Levels Are Inappropriately Elevated For The LevelNada MuchPas encore d'évaluation
- FCPS Oct 2015-1Document9 pagesFCPS Oct 2015-1Kahkashan HameedPas encore d'évaluation
- Management of Neonatal HypoglycemiaDocument14 pagesManagement of Neonatal Hypoglycemiaece142Pas encore d'évaluation
- ICD 11 CDDR Neurodevelopmental Disorders 2022 03 31Document70 pagesICD 11 CDDR Neurodevelopmental Disorders 2022 03 31apurbanmPas encore d'évaluation
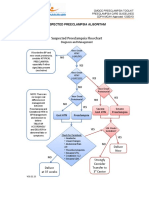
- Suspected Preeclampsia AlgorithmDocument1 pageSuspected Preeclampsia Algorithmmisstina.19876007100% (1)
- Group B1 (Cholera)Document70 pagesGroup B1 (Cholera)Krisianne Mae Lorenzo FranciscoPas encore d'évaluation
- Medical Nutrition Therapy For Cancer PreventionDocument5 pagesMedical Nutrition Therapy For Cancer PreventionotaPas encore d'évaluation
- DDEX37 Herald of The MoonDocument39 pagesDDEX37 Herald of The MoonCarl Max Bobo100% (5)
- Disertasi Dr. TerawanDocument8 pagesDisertasi Dr. TerawanNurhaidah AchmadPas encore d'évaluation
- High - Risk PregnancyDocument110 pagesHigh - Risk PregnancyAndre DityaPas encore d'évaluation
- Respiratory Distress of The New BornDocument15 pagesRespiratory Distress of The New BornJicko Street HooligansPas encore d'évaluation