Vous aimerez peut-être aussi
- BaruDocument11 pagesBaruJoelma Rabelo de PáduaPas encore d'évaluation
- Development and Characterization of DNA Microsatellite Primers For Buriti (Mauritia Flexuosa L.F.)Document5 pagesDevelopment and Characterization of DNA Microsatellite Primers For Buriti (Mauritia Flexuosa L.F.)vitocho_Pas encore d'évaluation
- Genetic Variability of Two Populations of Pseudoplatystoma Reticulatum From The Upper Paraguay River BasinDocument6 pagesGenetic Variability of Two Populations of Pseudoplatystoma Reticulatum From The Upper Paraguay River BasinOrlando Karim Shiro Jr.Pas encore d'évaluation
- Taxonomy and Conservation: A Case Study From Chamaedorea AlternansDocument9 pagesTaxonomy and Conservation: A Case Study From Chamaedorea AlternansumarPas encore d'évaluation
- Artigo Sobre STRDocument14 pagesArtigo Sobre STRapi-19990354Pas encore d'évaluation
- Genetic Structuring at A Fine Scale in The Russet Crowned Motmot (Momotus Mexicanus) in A Tropical Dry Forest in Central MexicoDocument4 pagesGenetic Structuring at A Fine Scale in The Russet Crowned Motmot (Momotus Mexicanus) in A Tropical Dry Forest in Central Mexicomadonna´sPas encore d'évaluation
- Development and Characterization of Microsatellite Markers For The Tropical Tree Species (Bignoniaceae)Document4 pagesDevelopment and Characterization of Microsatellite Markers For The Tropical Tree Species (Bignoniaceae)Vanessa BernardesPas encore d'évaluation
- 284 FullDocument8 pages284 FullKarlina SyahruddinPas encore d'évaluation
- Kohlmann2017 Stellatus MicrosDocument5 pagesKohlmann2017 Stellatus MicrosElena TaflanPas encore d'évaluation
- Alonso Etal 2014 Indv Variation Fecundity MethylationDocument5 pagesAlonso Etal 2014 Indv Variation Fecundity MethylationDianita PatitoPas encore d'évaluation
- Heterogeneous Genetic Structure in A Natural Population of Raulí (Nothofagus Nervosa)Document12 pagesHeterogeneous Genetic Structure in A Natural Population of Raulí (Nothofagus Nervosa)Luis Letelier GalvezPas encore d'évaluation
- Managing Habitat For The Eastern Tiger Salamander and Other Ambystomid Salamander SpeciesDocument45 pagesManaging Habitat For The Eastern Tiger Salamander and Other Ambystomid Salamander SpeciesEvelinaPas encore d'évaluation
- Smith 2016Document12 pagesSmith 2016karla rodriguezPas encore d'évaluation
- 1 Articulo The Population Genetic Diversity and Pattern of Pteroceltis Tatarinowii, A Relic Tree Endemic To China, Inferred From SSR MarkersDocument9 pages1 Articulo The Population Genetic Diversity and Pattern of Pteroceltis Tatarinowii, A Relic Tree Endemic To China, Inferred From SSR MarkersLuisa SuárezPas encore d'évaluation
- Villanueva 2015Document17 pagesVillanueva 2015Yerson TorresPas encore d'évaluation
- Issn: Issn:: Tillandsia LandbeckiiDocument13 pagesIssn: Issn:: Tillandsia LandbeckiiAnthony PaucaPas encore d'évaluation
- Colgan Et Al Manuscript For Submission MbeDocument21 pagesColgan Et Al Manuscript For Submission MbeywurmPas encore d'évaluation
- Pecoraro 2015Document6 pagesPecoraro 2015Fiy Jannatin AliyahPas encore d'évaluation
- Artigo - Ana Maria Olivatti - 2011Document6 pagesArtigo - Ana Maria Olivatti - 2011sabrinavfc1203Pas encore d'évaluation
- Phylogeny and Evolution of Basils and Allies (Ocimeae, Labiatae) Based On Three Plastid DNA RegionsDocument23 pagesPhylogeny and Evolution of Basils and Allies (Ocimeae, Labiatae) Based On Three Plastid DNA RegionslinubinoiPas encore d'évaluation
- POPULATION GENETICS OF Atta Sexdens Rubropilosa (Hymenoptera: Formicidae)Document16 pagesPOPULATION GENETICS OF Atta Sexdens Rubropilosa (Hymenoptera: Formicidae)Jose Fuentes MarioPas encore d'évaluation
- Check - Rapposa - Highly Conserved D-Loop Sequences in Woolly Mouse Opossums MarmosaDocument8 pagesCheck - Rapposa - Highly Conserved D-Loop Sequences in Woolly Mouse Opossums Marmosagbitencourt.bioPas encore d'évaluation
- Characterization of Microsatellite Loci of Tetragonisca AngustulaDocument5 pagesCharacterization of Microsatellite Loci of Tetragonisca AngustulaFabiana PiokerPas encore d'évaluation
- Jurnal Panduan Wajib DibacaDocument8 pagesJurnal Panduan Wajib DibacaEka NofiantiPas encore d'évaluation
- 10 26650-tjbc 1327318-3263743Document14 pages10 26650-tjbc 1327318-3263743Demet BiltekinPas encore d'évaluation
- ScirtothripsPhylogenyAnn Ent Soc Am2008Document11 pagesScirtothripsPhylogenyAnn Ent Soc Am2008ALONSOPas encore d'évaluation
- Contaminación de Los Suelos Con Huevos de Toxocara Canis en ParquesDocument12 pagesContaminación de Los Suelos Con Huevos de Toxocara Canis en ParquesJahir QHPas encore d'évaluation
- Genetic Linkage Map of A Cichlid Fish The Tilapia Oreochromis NiloticusDocument8 pagesGenetic Linkage Map of A Cichlid Fish The Tilapia Oreochromis Niloticusluis ruperto floresPas encore d'évaluation
- A Sequence Database For The Identification of Ectomycorrhizal Basidiomycetes by Phylogenetic AnalysisDocument16 pagesA Sequence Database For The Identification of Ectomycorrhizal Basidiomycetes by Phylogenetic Analysismayorca2310Pas encore d'évaluation
- CitriDocument4 pagesCitriAnonymous RkI7MG6gePas encore d'évaluation
- Cabrera Et Al. 2008 Araceae PhylogeneticsDocument13 pagesCabrera Et Al. 2008 Araceae PhylogeneticsGerardo A. SalazarPas encore d'évaluation
- Cap 1 Parasitic NematodeDocument24 pagesCap 1 Parasitic NematodeJorge Chavez FloresPas encore d'évaluation
- Zootaxa: Austrolebias Reicherti Loureiro & García, A Valid Species of Annual FishDocument15 pagesZootaxa: Austrolebias Reicherti Loureiro & García, A Valid Species of Annual FishFernando NunesPas encore d'évaluation
- Habitat Use by Sympatric Red and Roe Deer in A Mediterranean EcosystemDocument16 pagesHabitat Use by Sympatric Red and Roe Deer in A Mediterranean EcosystemAnindyaMustikaPas encore d'évaluation
- Genome Sizes in Diploid and Allopolyploid Arachis L. SpeciesDocument18 pagesGenome Sizes in Diploid and Allopolyploid Arachis L. SpeciesSofi VazquezPas encore d'évaluation
- Morphology, Reproductive Biology and Diet of Colubridae) in Brazilian AmazonDocument7 pagesMorphology, Reproductive Biology and Diet of Colubridae) in Brazilian AmazonEdiegoBatistaPas encore d'évaluation
- The Landscape of Human STR VariationDocument12 pagesThe Landscape of Human STR VariationJuliana SiruffoPas encore d'évaluation
- Mapping of Shoot y Tolerance Loci in Sorghum Using SSR MarkersDocument8 pagesMapping of Shoot y Tolerance Loci in Sorghum Using SSR MarkersBrent LagarizaPas encore d'évaluation
- BMC Evolutionary BiologyDocument15 pagesBMC Evolutionary BiologyAndrea ZuccoloPas encore d'évaluation
- PlatDocument6 pagesPlatFulano Mengano Sutano SerranoPas encore d'évaluation
- Meats, A. 2007Document11 pagesMeats, A. 2007Asriani NasirPas encore d'évaluation
- Thraustochtrid LyonsDocument7 pagesThraustochtrid LyonsdaneshnedaiePas encore d'évaluation
- Avise 1992 MolBiolEvolDocument17 pagesAvise 1992 MolBiolEvolIna Mae SisonPas encore d'évaluation
- Diversidad Gen. CedrelaDocument8 pagesDiversidad Gen. CedrelaJorge Luis Contreras HerreraPas encore d'évaluation
- BypassDocument5 pagesBypassGeorgina HernandezPas encore d'évaluation
- Gutierrez Liberatoetal2021Document13 pagesGutierrez Liberatoetal2021OSCAR ANDRES RODRIGUEZ FANDIÑOPas encore d'évaluation
- 2013 - SILVA Et Al. - New Species of GeastrumDocument12 pages2013 - SILVA Et Al. - New Species of GeastrumAna Clarissa RodriguesPas encore d'évaluation
- Metaphylogeny of 82 Gene Families Sheds A New Light On Chordate EvolutionDocument6 pagesMetaphylogeny of 82 Gene Families Sheds A New Light On Chordate EvolutionSatrio Haryu WPas encore d'évaluation
- 1 s2.0 S1470160X20302788 AmDocument38 pages1 s2.0 S1470160X20302788 AmZozoPas encore d'évaluation
- Microsatellite Markers Linked To QTL For Resistance To Mal de Rı o Cuarto Disease in Zea Mays LDocument7 pagesMicrosatellite Markers Linked To QTL For Resistance To Mal de Rı o Cuarto Disease in Zea Mays LmxdalloPas encore d'évaluation
- Distribution of Introns in The Mitochondrial Gene Nad1 in Land Plants: Phylogenetic and Molecular Evolutionary ImplicationsDocument18 pagesDistribution of Introns in The Mitochondrial Gene Nad1 in Land Plants: Phylogenetic and Molecular Evolutionary ImplicationslinubinoiPas encore d'évaluation
- Sanou 2015Document11 pagesSanou 2015José BlancasPas encore d'évaluation
- Pub - Encyclopedia of Genetics Genomics Proteomics and B PDFDocument4 046 pagesPub - Encyclopedia of Genetics Genomics Proteomics and B PDFraj252000100% (1)
- DNA Barcoding and Metabarcoding of Standardized Samples Reveal Patterns of Marine Benthic DiversityDocument17 pagesDNA Barcoding and Metabarcoding of Standardized Samples Reveal Patterns of Marine Benthic DiversityJennifer SilvaPas encore d'évaluation
- Ruiz-García Et Al 2021 Calbifrons, Potos, NasuaDocument48 pagesRuiz-García Et Al 2021 Calbifrons, Potos, NasuaManuel RuizPas encore d'évaluation
- Robe 2010 Amd DDCDocument12 pagesRobe 2010 Amd DDCGilberto Cavalheiro VieiraPas encore d'évaluation
- Artigo 7 - Biotic Potencial and Reproductive Parameters of Spodoptera Eridania (Stoll) (Lepidoptera, Noctuidae) in The LaboratoryDocument7 pagesArtigo 7 - Biotic Potencial and Reproductive Parameters of Spodoptera Eridania (Stoll) (Lepidoptera, Noctuidae) in The Laboratoryjean alex thimotheePas encore d'évaluation
- Diosque Et Al - 2003Document7 pagesDiosque Et Al - 2003oséias martins magalhãesPas encore d'évaluation
- Zaracho Et Al 2018Document13 pagesZaracho Et Al 2018amandavarago021Pas encore d'évaluation
- Collective Memory Conceptual Foundations and Theoretical Approaches PDFDocument10 pagesCollective Memory Conceptual Foundations and Theoretical Approaches PDFDaniel González EricesPas encore d'évaluation
- Framing The Apocalypse The PerformanceDocument25 pagesFraming The Apocalypse The PerformanceDaniel González EricesPas encore d'évaluation
- (15743012 - Religion and Theology) He Darkens Me With Brightness - The Theology of Pseudo-Dionysius in Hilduin's Vita of Saint Denis PDFDocument30 pages(15743012 - Religion and Theology) He Darkens Me With Brightness - The Theology of Pseudo-Dionysius in Hilduin's Vita of Saint Denis PDFDaniel González EricesPas encore d'évaluation
- (15743012 - Religion and Theology) He Darkens Me With Brightness - The Theology of Pseudo-Dionysius in Hilduin's Vita of Saint Denis PDFDocument30 pages(15743012 - Religion and Theology) He Darkens Me With Brightness - The Theology of Pseudo-Dionysius in Hilduin's Vita of Saint Denis PDFDaniel González EricesPas encore d'évaluation
- Collective Memory Conceptual Foundations and Theoretical Approaches PDFDocument10 pagesCollective Memory Conceptual Foundations and Theoretical Approaches PDFDaniel González EricesPas encore d'évaluation
- Collective Memory Conceptual Foundations and Theoretical Approaches PDFDocument10 pagesCollective Memory Conceptual Foundations and Theoretical Approaches PDFDaniel González EricesPas encore d'évaluation
- Beal A Dictionary of English Manuscript Terminology 1450 To 2000 2008Document478 pagesBeal A Dictionary of English Manuscript Terminology 1450 To 2000 2008ArcanaPas encore d'évaluation
- Value of Gold in Ancient EgyptDocument8 pagesValue of Gold in Ancient EgyptDaniel González EricesPas encore d'évaluation
- Ben Davies, Jana Funke - Sex, Gender and Time in Fiction and Culture-Palgrave Macmillan (2011) PDFDocument235 pagesBen Davies, Jana Funke - Sex, Gender and Time in Fiction and Culture-Palgrave Macmillan (2011) PDFDaniel González EricesPas encore d'évaluation
- Hans Van Meegeren, Forger: David Bratton, Virginia Benz, Emily Cook, & Barbara JohnsonDocument16 pagesHans Van Meegeren, Forger: David Bratton, Virginia Benz, Emily Cook, & Barbara JohnsonDaniel González EricesPas encore d'évaluation
- Meegeren 2Document4 pagesMeegeren 2Daniel González EricesPas encore d'évaluation
- Validez y Confiabilidad de La Versión C... Disorders Identification Test (AUDIT)Document10 pagesValidez y Confiabilidad de La Versión C... Disorders Identification Test (AUDIT)Daniel González EricesPas encore d'évaluation
- 23416364Document22 pages23416364Daniel González EricesPas encore d'évaluation
- Hans Van Meegeren, Forger: David Bratton, Virginia Benz, Emily Cook, & Barbara JohnsonDocument16 pagesHans Van Meegeren, Forger: David Bratton, Virginia Benz, Emily Cook, & Barbara JohnsonDaniel González EricesPas encore d'évaluation
- Indiana University Press Transactions of The Charles S. Peirce SocietyDocument28 pagesIndiana University Press Transactions of The Charles S. Peirce SocietyDaniel González EricesPas encore d'évaluation
- Pious Voices Nun-Scribes and The LanguagDocument32 pagesPious Voices Nun-Scribes and The LanguagDaniel González EricesPas encore d'évaluation
- (Paul Bywaters, Eileen McLeod) Social Work, Health (BookFi)Document226 pages(Paul Bywaters, Eileen McLeod) Social Work, Health (BookFi)Daniel González EricesPas encore d'évaluation
- Indiana University Press Transactions of The Charles S. Peirce SocietyDocument28 pagesIndiana University Press Transactions of The Charles S. Peirce SocietyDaniel González EricesPas encore d'évaluation
- S 0028688500012558Document9 pagesS 0028688500012558Daniel González EricesPas encore d'évaluation
- The Authority of The Word in St. John's Gospel: Charismatic Speech, Narrative Text, Logocentric MetaphysicsDocument24 pagesThe Authority of The Word in St. John's Gospel: Charismatic Speech, Narrative Text, Logocentric MetaphysicsDaniel González EricesPas encore d'évaluation
- MemraDocument3 pagesMemraDaniel González EricesPas encore d'évaluation
- Judith Butler, Gayatri Chakravorty Spivak Who Sings The Nation-State Language, Politics, BelongingDocument123 pagesJudith Butler, Gayatri Chakravorty Spivak Who Sings The Nation-State Language, Politics, Belongingolgi5407100% (3)
- Ccmed 0007-9731 2006 Num 49 195 2941Document40 pagesCcmed 0007-9731 2006 Num 49 195 2941Daniel González EricesPas encore d'évaluation
- 004057360306000205Document15 pages004057360306000205Daniel González EricesPas encore d'évaluation
- 3262920Document30 pages3262920Daniel González EricesPas encore d'évaluation
- An Interview With Peter H. Buschang PDFDocument11 pagesAn Interview With Peter H. Buschang PDFDaniel González EricesPas encore d'évaluation
- Tm0584 DescriptionDocument11 pagesTm0584 DescriptionDaniel González EricesPas encore d'évaluation
- Vista Previa de "Venetian Views, Venetian... F Venice - Google Books"Document1 pageVista Previa de "Venetian Views, Venetian... F Venice - Google Books"Daniel González EricesPas encore d'évaluation
- Vista Previa de "Racine - Language, Violen... y Reilly - Google Books"Document1 pageVista Previa de "Racine - Language, Violen... y Reilly - Google Books"Daniel González EricesPas encore d'évaluation
- Vista Previa de "Nowhere Is Perfect - Fren... Ystopias - Google Books"Document1 pageVista Previa de "Nowhere Is Perfect - Fren... Ystopias - Google Books"Daniel González EricesPas encore d'évaluation
- Toxicity of Clay Occulation of The Toxic Dino Agellate, Karenia Brevis, To Estuarine Invertebrates and SHDocument12 pagesToxicity of Clay Occulation of The Toxic Dino Agellate, Karenia Brevis, To Estuarine Invertebrates and SHinoussePas encore d'évaluation
- Earth and Life Science Week 14 (Perpetuation of Life) : DECEMBER 1-4, 2020Document9 pagesEarth and Life Science Week 14 (Perpetuation of Life) : DECEMBER 1-4, 2020Ann Kyla ManrezaPas encore d'évaluation
- Guia de Algas Bocas Del ToroDocument71 pagesGuia de Algas Bocas Del ToroNohemi PaolaPas encore d'évaluation
- Points To Remember: Morphology of Flowering PlantsDocument8 pagesPoints To Remember: Morphology of Flowering PlantsMuskan VarlaniPas encore d'évaluation
- Mammoth KillDocument2 pagesMammoth KillLê Thị Thanh ThảoPas encore d'évaluation
- ProteinDocument23 pagesProteinsamantha garciaPas encore d'évaluation
- Barquín Et Al., 2011.Document18 pagesBarquín Et Al., 2011.D'imrPoloPas encore d'évaluation
- Lesson-Plan Ecologica RelationshipDocument10 pagesLesson-Plan Ecologica RelationshipRen Mart Decipeda100% (3)
- Terra Rosa Emag #20Document61 pagesTerra Rosa Emag #20Terra Rosa100% (1)
- NParks GDP Guidelines Version 3Document214 pagesNParks GDP Guidelines Version 3Lim Kang HaiPas encore d'évaluation
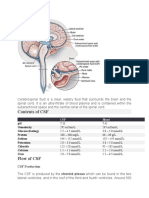
- Cerebrospinal FluidDocument3 pagesCerebrospinal FluidlecturioPas encore d'évaluation
- Goby Shirmp MutualismDocument20 pagesGoby Shirmp MutualismKhandoker Raisul AzadPas encore d'évaluation
- Big Six PDFDocument4 pagesBig Six PDFmotibaPas encore d'évaluation
- Unesco - Eolss Sample Chapters: History of BiologyDocument6 pagesUnesco - Eolss Sample Chapters: History of BiologyLauraLópezGiraldoPas encore d'évaluation
- Article: Contribution To The Knowledge of Ephemeroptera (Insecta) From Goiás State, BrazilDocument15 pagesArticle: Contribution To The Knowledge of Ephemeroptera (Insecta) From Goiás State, BrazilErikcsen Augusto RaimundiPas encore d'évaluation
- MD II Material Pentru 24.03.2020Document5 pagesMD II Material Pentru 24.03.2020Irina Panciu StefanPas encore d'évaluation
- EXERCISE 4 Stains PDFDocument3 pagesEXERCISE 4 Stains PDFOsannah Irish InsongPas encore d'évaluation
- Review of LaboratoryDocument48 pagesReview of LaboratoryIkhar RidhoPas encore d'évaluation
- Anatomical Features of Bougainvillea (Nyctaginaceae) : Sarah ChewDocument7 pagesAnatomical Features of Bougainvillea (Nyctaginaceae) : Sarah ChewPratistha ShauryaPas encore d'évaluation
- ANIMILIADocument10 pagesANIMILIAAnand Teekmani AnuPas encore d'évaluation
- Nooij18 Frontiers OverviewVirusMetagenomicClassificationMethodsDocument21 pagesNooij18 Frontiers OverviewVirusMetagenomicClassificationMethodsAnh H NguyenPas encore d'évaluation
- IV Fluid Management: Islam Awni Abu SamraDocument41 pagesIV Fluid Management: Islam Awni Abu SamraIslam AwniPas encore d'évaluation
- Microbiology 1.03 Basic Concepts 1Document9 pagesMicrobiology 1.03 Basic Concepts 1Camila BarzagaPas encore d'évaluation
- Contruccion de Vector de Expresion y Evaluación de Rendimiento de Biosimilar de Rituximab en CHODocument7 pagesContruccion de Vector de Expresion y Evaluación de Rendimiento de Biosimilar de Rituximab en CHOSebastián Zapata LoaizaPas encore d'évaluation
- Unit Ii Biomedical Perspective in Gender and SexualityDocument46 pagesUnit Ii Biomedical Perspective in Gender and SexualityRowena Lanete ButaPas encore d'évaluation
- Esthetic Dentistry / Orthodontic Courses by Indian Dental AcademyDocument256 pagesEsthetic Dentistry / Orthodontic Courses by Indian Dental Academyindian dental academyPas encore d'évaluation
- CBSE Class 12 Biology Marking Scheme and Answer Key Term 1 2021 22Document4 pagesCBSE Class 12 Biology Marking Scheme and Answer Key Term 1 2021 22Prashanth 070Pas encore d'évaluation
- Thesis - DraftDocument45 pagesThesis - DraftFlora Fauna FlavourPas encore d'évaluation
- Microsoft PowerPoint - Defense Mechanisms of The Gingiva (Compatibility MDocument19 pagesMicrosoft PowerPoint - Defense Mechanisms of The Gingiva (Compatibility MDon Vigis AbrahamPas encore d'évaluation