Vous aimerez peut-être aussi
- In Defense of Processed FoodDocument279 pagesIn Defense of Processed FoodАлексей ШишкинPas encore d'évaluation
- Vol. Biological Of: Value ProteinsDocument2 pagesVol. Biological Of: Value ProteinssdgwePas encore d'évaluation
- Biological Evaluation of Proteins A New AspectDocument9 pagesBiological Evaluation of Proteins A New AspectsdgwePas encore d'évaluation
- Am J Clin Nutr 1981 Potter 328 34Document7 pagesAm J Clin Nutr 1981 Potter 328 34Abhishek Rekha KoshtaPas encore d'évaluation
- Korelesk 182Document8 pagesKorelesk 182Vimal KumarPas encore d'évaluation
- Aleator 2000 Low Protein Amino Acid Supplemented Diets in Broiler Chickens, Effects On Performance, Carcass Characteristics, Whole Body Composition and Efficiencie of Nutrient UtilisationDocument8 pagesAleator 2000 Low Protein Amino Acid Supplemented Diets in Broiler Chickens, Effects On Performance, Carcass Characteristics, Whole Body Composition and Efficiencie of Nutrient UtilisationMiguelAngelMatusAragonPas encore d'évaluation
- Effect of A Hyper-Protein Diet On Wistar Rats Development and Intestinal FunctionDocument6 pagesEffect of A Hyper-Protein Diet On Wistar Rats Development and Intestinal FunctionhafizPas encore d'évaluation
- The Effect of Fibrolytic Exogenous Enzyme On Fattening Performance of SteersDocument6 pagesThe Effect of Fibrolytic Exogenous Enzyme On Fattening Performance of SteersMuhammad Khairul RaisPas encore d'évaluation
- Lec 8 Protein QualityDocument6 pagesLec 8 Protein QualityABISAGANA B CS&BS studentPas encore d'évaluation
- Note On The Determination of Net Protein Utilization by Carcass AnalysisDocument2 pagesNote On The Determination of Net Protein Utilization by Carcass AnalysisCesar OliveiraPas encore d'évaluation
- Nonruminant Nutrition: Amino Acids 1Document3 pagesNonruminant Nutrition: Amino Acids 1Rangga AlloysPas encore d'évaluation
- Bioavailability of Raw and Cooked EggsDocument7 pagesBioavailability of Raw and Cooked EggsVirtuosoXPas encore d'évaluation
- Dietary Protein Effect On Parakeet ReproductionDocument6 pagesDietary Protein Effect On Parakeet ReproductionwalriePas encore d'évaluation
- OK - Effect of Dietary Butyric Acid SupplementationDocument8 pagesOK - Effect of Dietary Butyric Acid SupplementationOliver TalipPas encore d'évaluation
- Animals 03 00558Document16 pagesAnimals 03 00558SAADPas encore d'évaluation
- Comparacion de Energia Metabolizable y Energia Productiva en Pollos en CrecimientoDocument17 pagesComparacion de Energia Metabolizable y Energia Productiva en Pollos en CrecimientoDanny ProañoPas encore d'évaluation
- NutritionDocument21 pagesNutritionAnanthasetty NagadurgaPas encore d'évaluation
- Ostrich Meat Protein Quality and DigestibilityDocument5 pagesOstrich Meat Protein Quality and DigestibilitysuperboysingurPas encore d'évaluation
- The Effect of Synbiotic On Carcass Percentage and Abdominal Fat Percentage of Native ChickenDocument5 pagesThe Effect of Synbiotic On Carcass Percentage and Abdominal Fat Percentage of Native ChickenIJAR JOURNALPas encore d'évaluation
- The Efficacy of A Phytase Enzyme Fed To Hy-Line WDocument7 pagesThe Efficacy of A Phytase Enzyme Fed To Hy-Line Wdanielleayyash1Pas encore d'évaluation
- Effect of Betaine Supplementation To Methionine-Deficient Diet On Growth Performance, Carcass Characteristics, Blood Parameters and Economic Efficiency of BroilersDocument10 pagesEffect of Betaine Supplementation To Methionine-Deficient Diet On Growth Performance, Carcass Characteristics, Blood Parameters and Economic Efficiency of BroilersSusmita ThullimalliPas encore d'évaluation
- Vitamin eDocument7 pagesVitamin eJonathan ShaoPas encore d'évaluation
- Methods of Estimating Protein Quality: Biological Value (BV)Document20 pagesMethods of Estimating Protein Quality: Biological Value (BV)Nguyen PhamPas encore d'évaluation
- Improving Phosphorus Availability in Broiler Diets Based On Corn-Soybean Meal Using Microbial PhytaseDocument7 pagesImproving Phosphorus Availability in Broiler Diets Based On Corn-Soybean Meal Using Microbial PhytaseSarah Tarala MoscosaPas encore d'évaluation
- Track 3 Eating Patterns and Behaviour P63Document15 pagesTrack 3 Eating Patterns and Behaviour P63Yuriko AndrePas encore d'évaluation
- Estimation of The Biological Value of Food Proteins by A Modified Equation of The Essential Amino Acid Index and The Chemical ScoreDocument6 pagesEstimation of The Biological Value of Food Proteins by A Modified Equation of The Essential Amino Acid Index and The Chemical ScoreCourtney KayPas encore d'évaluation
- Jurnal Onrum WidoDocument4 pagesJurnal Onrum WidoBiokta WahyudiPas encore d'évaluation
- Jansman 2002Document12 pagesJansman 2002Yeonjun ChoiPas encore d'évaluation
- Asad Abbas 2019-Ag-8718 - Measures of Protein Quality For Mono-GastricsDocument12 pagesAsad Abbas 2019-Ag-8718 - Measures of Protein Quality For Mono-GastricsAsad AbbasPas encore d'évaluation
- Physiology of Metabolism and EnergyDocument44 pagesPhysiology of Metabolism and EnergyShilvia JungPas encore d'évaluation
- Relationship To Muscle Triglyceride and w-3 Fatty Acids in Muscle PhospholipidDocument10 pagesRelationship To Muscle Triglyceride and w-3 Fatty Acids in Muscle PhospholipidAdeess AengPas encore d'évaluation
- Metabolizable Energy and Digestible Amino Acids of Full-Fat Soybean Without or With Protease Supplementation in Diets For BroilersDocument12 pagesMetabolizable Energy and Digestible Amino Acids of Full-Fat Soybean Without or With Protease Supplementation in Diets For BroilerssanielPas encore d'évaluation
- Poultry Research Report 29: I N T R o D U C T I o NDocument4 pagesPoultry Research Report 29: I N T R o D U C T I o NRucha ZombadePas encore d'évaluation
- 1 s2.0 S0022030267876161 MainDocument5 pages1 s2.0 S0022030267876161 MainFebri Daris Faidatur RohmahPas encore d'évaluation
- Perhitungan PDFDocument9 pagesPerhitungan PDFYusep SismaryantoPas encore d'évaluation
- Pengaruh Penggunaan Ajitein Dalam Pakan Terhadap Produksi Dan Kualitas Susu Sapi PerahDocument10 pagesPengaruh Penggunaan Ajitein Dalam Pakan Terhadap Produksi Dan Kualitas Susu Sapi PerahsuyitnoPas encore d'évaluation
- Vanvliet 2017Document12 pagesVanvliet 2017Adrián MartínPas encore d'évaluation
- Dietary Protein Oscillation Effects On Feed IntakeDocument13 pagesDietary Protein Oscillation Effects On Feed IntakeParaschiva PîțuPas encore d'évaluation
- The United Republic of Tanzania Ministry of Livestock and Fisheries Development Fisheries Education and Training AgencyDocument7 pagesThe United Republic of Tanzania Ministry of Livestock and Fisheries Development Fisheries Education and Training AgencyOG AdamPas encore d'évaluation
- VacunosDocument10 pagesVacunosLeoCallePas encore d'évaluation
- P. Styli-P. Uannamei P. Uannamei P. Setiferus P. AztecusDocument12 pagesP. Styli-P. Uannamei P. Uannamei P. Setiferus P. AztecusBruna Paula Torres QuintoPas encore d'évaluation
- 35 Eggs Per DayDocument4 pages35 Eggs Per DayVlad GubernatPas encore d'évaluation
- 01 1159.fullDocument12 pages01 1159.fullCiandeb GracianoPas encore d'évaluation
- O. Colin-Schoellen, S. Jurjanz, F. Laurent: Laboratoire de Sciences AnimalesDocument13 pagesO. Colin-Schoellen, S. Jurjanz, F. Laurent: Laboratoire de Sciences AnimalesOmary2jPas encore d'évaluation
- Dietary Choline For The Lactating Cow. Possible Effects On Milk Fat Synthesis - Erdam, Shaver y VandersallDocument6 pagesDietary Choline For The Lactating Cow. Possible Effects On Milk Fat Synthesis - Erdam, Shaver y Vandersalljamonto5Pas encore d'évaluation
- Dietary Protein or Energy Restriction Influences Age and Weight at Puberty in Ewes Lambs Boulanouar Et Al 1995Document10 pagesDietary Protein or Energy Restriction Influences Age and Weight at Puberty in Ewes Lambs Boulanouar Et Al 1995Canuto MunozPas encore d'évaluation
- Reduction in Dietary Lysine Increases Muscle Free Amino Acids Through Changes in Protein Metabolism in ChickensDocument9 pagesReduction in Dietary Lysine Increases Muscle Free Amino Acids Through Changes in Protein Metabolism in Chickensalex stefanutPas encore d'évaluation
- Pengaruh Penggunaan Ajitein Dalam Pakan Terhadap Produksi Dan Kualitas Susu Sapi PerahDocument10 pagesPengaruh Penggunaan Ajitein Dalam Pakan Terhadap Produksi Dan Kualitas Susu Sapi PerahVera AlfianiPas encore d'évaluation
- Biological ValueDocument4 pagesBiological ValueKedar Singh ThakuriPas encore d'évaluation
- Effect of Feeding On Growth and Blood Biochemistry of Male Fallow DeerDocument3 pagesEffect of Feeding On Growth and Blood Biochemistry of Male Fallow Deer陳克論Pas encore d'évaluation
- ACMS Import Proteina e TF Na HipetrofiaDocument5 pagesACMS Import Proteina e TF Na HipetrofiaWellington Fernando da SilvaPas encore d'évaluation
- Levels of Lysine and Methionine+cystine For Growing New Zealand White RabbitsDocument34 pagesLevels of Lysine and Methionine+cystine For Growing New Zealand White RabbitsRogueCheneyPas encore d'évaluation
- Pittroff 2006Document16 pagesPittroff 2006MellianyPas encore d'évaluation
- Vandenbraak2013 - CoagulaciónDocument7 pagesVandenbraak2013 - CoagulaciónLeti PeraltaPas encore d'évaluation
- Effect of Exogenous Enzymes in Maize Based Diets Varying in Nutrient Density For Young Broilers Growth Performance and Digestibility of EnergyDocument9 pagesEffect of Exogenous Enzymes in Maize Based Diets Varying in Nutrient Density For Young Broilers Growth Performance and Digestibility of Energyshahbaz zafarPas encore d'évaluation
- Acta Veterinaria (Beograd), Vol. 59, No. 5-6, 535-545, 2009Document11 pagesActa Veterinaria (Beograd), Vol. 59, No. 5-6, 535-545, 2009Ngoc NguyenPas encore d'évaluation
- Bi 9740067Document8 pagesBi 9740067Bachi VarmaPas encore d'évaluation
- Diet in Indian Diabetics: Nutrition SectionDocument8 pagesDiet in Indian Diabetics: Nutrition Sectionlaloo01Pas encore d'évaluation
- Influence of Feeding Dehydrated Poultry Waste On Broiler Growth, and Meat Flavor, and CompositionDocument6 pagesInfluence of Feeding Dehydrated Poultry Waste On Broiler Growth, and Meat Flavor, and CompositionEz DoradoPas encore d'évaluation
- Nutrition Conference for Feed Manufacturers: University of Nottingham, Volume 7D'EverandNutrition Conference for Feed Manufacturers: University of Nottingham, Volume 7Pas encore d'évaluation
- Recent Advances in Animal Nutrition– 1978: Studies in the Agricultural and Food SciencesD'EverandRecent Advances in Animal Nutrition– 1978: Studies in the Agricultural and Food SciencesPas encore d'évaluation
- Stunting TreatmentDocument10 pagesStunting TreatmentDiana etika azzahraPas encore d'évaluation
- Malnutrition-Journal WDocument11 pagesMalnutrition-Journal Wgeng gengPas encore d'évaluation
- MKU Material Plus Daftar IsiDocument126 pagesMKU Material Plus Daftar Isitata aprianiPas encore d'évaluation
- SGC Concept Note - Project Brief Template 1Document4 pagesSGC Concept Note - Project Brief Template 1Shawie TabladaPas encore d'évaluation
- GPAF (IMP 056) Project Completion Report 31 May ChildHope UK 20170320060338Document54 pagesGPAF (IMP 056) Project Completion Report 31 May ChildHope UK 20170320060338Layla NdaulaPas encore d'évaluation
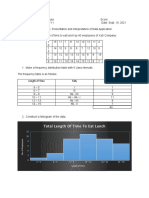
- Cedrix James Estoquia - OLLC Lesson 4.6 Presentation and Interpretation of Data ApplicationDocument4 pagesCedrix James Estoquia - OLLC Lesson 4.6 Presentation and Interpretation of Data ApplicationDeuK WRPas encore d'évaluation
- Ni Kadek Dwi Anjani - SICH 2020Document11 pagesNi Kadek Dwi Anjani - SICH 2020Dwie Anjani SaiPas encore d'évaluation
- Consumer BehaviourDocument11 pagesConsumer BehaviourNaman Agarwal [PG23]Pas encore d'évaluation
- Lpe 2501 - SCL 6Document8 pagesLpe 2501 - SCL 6NUR AQILLAH NATASHA BINTI SOBRI / UPMPas encore d'évaluation
- Composition of CarbohydratesDocument6 pagesComposition of Carbohydratesgeorgejhon1949Pas encore d'évaluation
- Fel Non PreggDocument1 pageFel Non PreggCLARISSE GEMROSE CUÑADAPas encore d'évaluation
- Sridevi Jagadeesan Dietetics Resume FinalDocument2 pagesSridevi Jagadeesan Dietetics Resume Finalapi-344829729Pas encore d'évaluation
- KONSELING GIZI PARIWISATA 2 OrgDocument6 pagesKONSELING GIZI PARIWISATA 2 OrgSintia AriyuniPas encore d'évaluation
- Development Pathways Toward Zero HungerDocument14 pagesDevelopment Pathways Toward Zero HungerSergio Andrés GonzálezPas encore d'évaluation
- Vitamin D PDFDocument23 pagesVitamin D PDFBroo RoorPas encore d'évaluation
- 19 Smartime 4 Test 4Document5 pages19 Smartime 4 Test 4Rodrigo MuñozPas encore d'évaluation
- Nursing Care Plan: Western Mindanao State University College of NursingDocument11 pagesNursing Care Plan: Western Mindanao State University College of NursingPatricia VasquezPas encore d'évaluation
- 10 Fat Extraction LabDocument2 pages10 Fat Extraction Labraanja2Pas encore d'évaluation
- Make X Do X LetDocument4 pagesMake X Do X LetSue B. RodriguesPas encore d'évaluation
- An Anti-Inflammatory Diet May Be Good For Your Joints - Harvard HealthDocument10 pagesAn Anti-Inflammatory Diet May Be Good For Your Joints - Harvard HealthAdil ShaikPas encore d'évaluation
- Chapter 1 Introduction To NutritionDocument55 pagesChapter 1 Introduction To Nutritionikmal hazimPas encore d'évaluation
- Form - Case Analysis Essay The Children of Sitio MabolonDocument1 pageForm - Case Analysis Essay The Children of Sitio MabolonTianboyPas encore d'évaluation
- Biochemical AssessmentDocument2 pagesBiochemical AssessmentRecca Maureen PabloPas encore d'évaluation
- Minutes of Meeting Sept 2, 2022Document11 pagesMinutes of Meeting Sept 2, 2022roseannurakPas encore d'évaluation
- Nature Activities Binder Variant PurpleDocument21 pagesNature Activities Binder Variant PurpleAngel Kate PolitudPas encore d'évaluation
- Changing Time of Feeding Starter Grower and FinishDocument9 pagesChanging Time of Feeding Starter Grower and FinishSurani RahmanPas encore d'évaluation
- Nutrion Counseling Project Nufd 482Document22 pagesNutrion Counseling Project Nufd 482api-608128291Pas encore d'évaluation
- WFP 0000141430Document2 pagesWFP 0000141430ELBOU ALY SIDINAPas encore d'évaluation
- CBYDP Brgy. Macabiso Sultan MasturaDocument4 pagesCBYDP Brgy. Macabiso Sultan MasturaJasmin Cabudoy BalabaganPas encore d'évaluation