Vous aimerez peut-être aussi
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (121)
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (588)
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (400)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (266)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5794)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2259)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (345)
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (895)
- Pruning and Training What When and How To Prune Revised EditionDocument338 pagesPruning and Training What When and How To Prune Revised EditionAlexandraLammerink100% (1)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (74)
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- 8.aviation, Space and Deep Sea Diving PhysiologyDocument5 pages8.aviation, Space and Deep Sea Diving PhysiologyCLEMENTPas encore d'évaluation
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- Mechanical VentilationDocument16 pagesMechanical VentilationArt Christian Ramos88% (8)
- Hema 2 - Prelim Topic 1-Intro To HemostasisDocument4 pagesHema 2 - Prelim Topic 1-Intro To HemostasisLowenstein JenzenPas encore d'évaluation
- NCPsDocument13 pagesNCPsRocel DevillesPas encore d'évaluation
- Msds Buffer AsetatDocument6 pagesMsds Buffer AsetatAnnisa SeptianiPas encore d'évaluation
- Lesson 3. Physical Properties of Biomaterials Like Shape, Size, Volume and Surface AreaDocument4 pagesLesson 3. Physical Properties of Biomaterials Like Shape, Size, Volume and Surface AreaAbhishekShuklaPas encore d'évaluation
- Syllabus NCM 103Document11 pagesSyllabus NCM 103ivanguzman86Pas encore d'évaluation
- DXN Is The Perfect BusinessDocument45 pagesDXN Is The Perfect Businesssomasekharvasudevan83% (6)
- This Column Is Reprinted, by Permission, From The National Psychologist, 2003Document1 pageThis Column Is Reprinted, by Permission, From The National Psychologist, 2003AbhishekShuklaPas encore d'évaluation
- Food Chemistry: M.W. Korir, F.N. Wachira, J.K. Wanyoko, R.M. Ngure, R. KhalidDocument1 pageFood Chemistry: M.W. Korir, F.N. Wachira, J.K. Wanyoko, R.M. Ngure, R. KhalidAbhishekShuklaPas encore d'évaluation
- Related Titles: (ISBN 978-1-84569-648-1) (ISBN 978-1-84569-983-3) (ISBN 978-1-85573-463-0)Document1 pageRelated Titles: (ISBN 978-1-84569-648-1) (ISBN 978-1-84569-983-3) (ISBN 978-1-85573-463-0)AbhishekShuklaPas encore d'évaluation
- Preface: Fereidoon ShahidiDocument1 pagePreface: Fereidoon ShahidiAbhishekShuklaPas encore d'évaluation
- JF 970948 LDocument5 pagesJF 970948 LAbhishekShuklaPas encore d'évaluation
- Class Time Table For January-April 2014: Department of Chemical Engineering Date: 30/12/2013Document2 pagesClass Time Table For January-April 2014: Department of Chemical Engineering Date: 30/12/2013AbhishekShuklaPas encore d'évaluation
- 1 KjuhygtDocument1 page1 KjuhygtAbhishekShuklaPas encore d'évaluation
- Supercritical Technology in Brazil: System Investigated (1994-2003)Document9 pagesSupercritical Technology in Brazil: System Investigated (1994-2003)AbhishekShuklaPas encore d'évaluation
- Food Chemistry: I. Stoilova, A. Krastanov, A. Stoyanova, P. Denev, S. GargovaDocument7 pagesFood Chemistry: I. Stoilova, A. Krastanov, A. Stoyanova, P. Denev, S. GargovaAbhishekShuklaPas encore d'évaluation
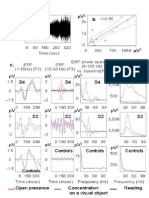
- Supplementary Figure 2: Modulation of The Event-Related Potential (ERP) by TheDocument1 pageSupplementary Figure 2: Modulation of The Event-Related Potential (ERP) by TheAbhishekShuklaPas encore d'évaluation
- Discussion: Long-Distance Gamma SynchronyDocument1 pageDiscussion: Long-Distance Gamma SynchronyAbhishekShuklaPas encore d'évaluation
- Supplementary Figure 5Document1 pageSupplementary Figure 5AbhishekShuklaPas encore d'évaluation
- Qbank FormatDocument5 pagesQbank FormatpashaPas encore d'évaluation
- 5090 w10 Ms 61Document3 pages5090 w10 Ms 61mstudy123456Pas encore d'évaluation
- General Biology 1: TEST II. Modified True or False. Indicate Whether The Statement Is True or False. If False, Change TheDocument2 pagesGeneral Biology 1: TEST II. Modified True or False. Indicate Whether The Statement Is True or False. If False, Change TheMA. HAZEL TEOLOGOPas encore d'évaluation
- Anti-Depressant Drugs: Presented by L.Nithish Shankar Ii Year Mbbs KGMCDocument17 pagesAnti-Depressant Drugs: Presented by L.Nithish Shankar Ii Year Mbbs KGMCÑiťhišh Śhankąŕ LóganáthánPas encore d'évaluation
- Exp 1 Group 8 MT3 ADocument12 pagesExp 1 Group 8 MT3 AJoanna RamosoPas encore d'évaluation
- UntitledDocument11 pagesUntitledZeianne BosabicaPas encore d'évaluation
- Ejpd 2014 4 13Document7 pagesEjpd 2014 4 13javi222222Pas encore d'évaluation
- Physioex Lab Report: Pre-Lab Quiz ResultsDocument3 pagesPhysioex Lab Report: Pre-Lab Quiz ResultsNicole de LeonPas encore d'évaluation
- PM Artifacts WEBDocument20 pagesPM Artifacts WEBSalman MemonPas encore d'évaluation
- PenisilinDocument11 pagesPenisilinHaruka KanataPas encore d'évaluation
- Detailed Lesson Plan in Endocrine SystemDocument4 pagesDetailed Lesson Plan in Endocrine Systemjonelllantero032Pas encore d'évaluation
- TEP 2, Thera Ex 1 and Physiology 2 SyllabusDocument2 pagesTEP 2, Thera Ex 1 and Physiology 2 SyllabusJohnpeter EsporlasPas encore d'évaluation
- 1ST Proff MBBS Papers (2021-11)Document66 pages1ST Proff MBBS Papers (2021-11)drbishalchowdhury13Pas encore d'évaluation
- Kristal Pada UrineDocument4 pagesKristal Pada UrineyholicPas encore d'évaluation
- Case Report Myoma Uterine: Supervised By: Dr. Hesty Duhita Permata, SP - OGDocument12 pagesCase Report Myoma Uterine: Supervised By: Dr. Hesty Duhita Permata, SP - OGNiki Rizqi rachmawatiPas encore d'évaluation
- 3 Pranayama Exercises For SingersDocument3 pages3 Pranayama Exercises For SingersGypsy SingerPas encore d'évaluation
- CPR Review Worksheet Ans KeyDocument9 pagesCPR Review Worksheet Ans Keyapi-234803876Pas encore d'évaluation
- Oral Lec 8Document9 pagesOral Lec 8Adam AliraqiPas encore d'évaluation
- ISC Class 11 Bio Specimen PaperDocument6 pagesISC Class 11 Bio Specimen PaperparamPas encore d'évaluation
- Anatomy and Physiology of Respiratory System Tutorial2Document1 pageAnatomy and Physiology of Respiratory System Tutorial2Jhullian Frederick Val VergaraPas encore d'évaluation
- 11 Brain & LanguageDocument20 pages11 Brain & LanguageIvan Giovanni MiterPas encore d'évaluation
- Perfusionist Job DescriptionDocument3 pagesPerfusionist Job DescriptionNouman IshaqPas encore d'évaluation