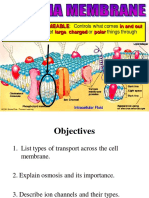
Vous aimerez peut-être aussi
- Active Transport MethodsDocument13 pagesActive Transport Methodssrikanth100% (3)
- 2222 TransportDocument42 pages2222 TransportaddwrPas encore d'évaluation
- Active Transport MechanismDocument46 pagesActive Transport MechanismnoblefxPas encore d'évaluation
- ACTIVE TRANSPORT Vs PASSIVE TRANSPORTDocument6 pagesACTIVE TRANSPORT Vs PASSIVE TRANSPORTfelixnellyPas encore d'évaluation
- Cell Membrane Transport Types & MechanismsDocument92 pagesCell Membrane Transport Types & MechanismsMohammed RamzyPas encore d'évaluation
- Assignment OneDocument6 pagesAssignment OneEmmanuel OpokuPas encore d'évaluation
- GC University Faisalabad Cell Biology Document on Membrane TransportDocument5 pagesGC University Faisalabad Cell Biology Document on Membrane TransportShahzad Ahmad NomanPas encore d'évaluation
- ACTIVE TRANSPORT MECHANISMS MOVE SUBSTANCES AGAINST GRADIENTSDocument10 pagesACTIVE TRANSPORT MECHANISMS MOVE SUBSTANCES AGAINST GRADIENTSSylvia DuzonPas encore d'évaluation
- Plant Cell Transport Proteins and Membrane StructuresDocument20 pagesPlant Cell Transport Proteins and Membrane StructuresBrandon BarndtPas encore d'évaluation
- Transport Across Membranes FinalDocument77 pagesTransport Across Membranes FinalLorraine Eve AbilaPas encore d'évaluation
- Active TransportDocument51 pagesActive Transportericka khimPas encore d'évaluation
- Drug Absorption-WPS OfficeDocument6 pagesDrug Absorption-WPS Officeloknath sur parthoPas encore d'évaluation
- This Energy Is Harvested From ATP That Is Generated Through Cellular Metabolism.)Document4 pagesThis Energy Is Harvested From ATP That Is Generated Through Cellular Metabolism.)AnnEvitePas encore d'évaluation
- ATP-Powered Active Transport Moves Molecules Against GradientsDocument4 pagesATP-Powered Active Transport Moves Molecules Against GradientsAnne NicolePas encore d'évaluation
- Cell Transport MechanismsDocument4 pagesCell Transport MechanismsAlan VarghesePas encore d'évaluation
- Practical Work 3 - Measuring Active Ion TransportDocument7 pagesPractical Work 3 - Measuring Active Ion TransportAchraf RabadiPas encore d'évaluation
- Transport 2 PDF Lecture Elalamein 2021-22Document5 pagesTransport 2 PDF Lecture Elalamein 2021-22Ahmed MagdyPas encore d'évaluation
- Nutrition EnergeticsDocument13 pagesNutrition EnergeticsLuis EduardoPas encore d'évaluation
- 2Q Week 4 Lesson 4 ChemiosmosisDocument13 pages2Q Week 4 Lesson 4 ChemiosmosisKC DinglasPas encore d'évaluation
- Plant Physiology Week 4Document68 pagesPlant Physiology Week 4PENELOPE LAWAGANPas encore d'évaluation
- Mastering Biology Section 94 FlashcardsDocument8 pagesMastering Biology Section 94 Flashcardsyoongkang0122Pas encore d'évaluation
- Biophysics-Lectures-2012-3 (Physiology) PDFDocument107 pagesBiophysics-Lectures-2012-3 (Physiology) PDFNedelcuGeorgePas encore d'évaluation
- Bio Notes FinalsDocument4 pagesBio Notes FinalsJoaquine ArateaPas encore d'évaluation
- Finals NotesDocument21 pagesFinals NotesJoaquine ArateaPas encore d'évaluation
- Active TransportDocument14 pagesActive TransportRjay CruzPas encore d'évaluation
- Biology Study StudyGuideDocument18 pagesBiology Study StudyGuideMelissaPas encore d'évaluation
- REVISION 3Document27 pagesREVISION 3Biology BảoPas encore d'évaluation
- Oxidative Phosphorylation - Biology (Article) - Khan Academy - 1623380772260Document19 pagesOxidative Phosphorylation - Biology (Article) - Khan Academy - 1623380772260Ismayer EfeapePas encore d'évaluation
- Chap 11 MbotcDocument4 pagesChap 11 MbotcĐồng Ngọc HàPas encore d'évaluation
- Note For Grade 10Document10 pagesNote For Grade 10peter20000.npkPas encore d'évaluation
- Bio physics internal energy and enthalpyDocument8 pagesBio physics internal energy and enthalpySabika HassanPas encore d'évaluation
- Chapter 7 Membrane Structure and FunctionDocument4 pagesChapter 7 Membrane Structure and FunctionMohammad Mahrus Bin AshfaqPas encore d'évaluation
- 1st Year 2nd Week PHYS Lecture 6 2019 2020Document14 pages1st Year 2nd Week PHYS Lecture 6 2019 2020Ahmed TarekPas encore d'évaluation
- Test Essays January 4Document7 pagesTest Essays January 4Stephanie JenkinsPas encore d'évaluation
- Active TransportDocument8 pagesActive TransportSarah NasutionPas encore d'évaluation
- Topic 3-Active Transport - EditedDocument7 pagesTopic 3-Active Transport - Editedsalve joy villanuevaPas encore d'évaluation
- BIOC 2061 Tutorial 3Document4 pagesBIOC 2061 Tutorial 3Kavita MaharajPas encore d'évaluation
- Key Points RevisionDocument124 pagesKey Points RevisionSumeyyah KemalPas encore d'évaluation
- 3 - kh201295 kh201295Document5 pages3 - kh201295 kh201295murtada gubaPas encore d'évaluation
- Channels and PumpsDocument10 pagesChannels and Pumpsazazel666Pas encore d'évaluation
- Active Vesicular TransportDocument14 pagesActive Vesicular Transportsarah abarcaPas encore d'évaluation
- Molecular Cell Biology Lodish 7th Edition Solutions ManualDocument8 pagesMolecular Cell Biology Lodish 7th Edition Solutions ManualKennethHendersonyecg100% (39)
- Homeostasis E (X) NdocytosisDocument18 pagesHomeostasis E (X) NdocytosisJeff ParkPas encore d'évaluation
- Cell Membrane Transport: Active, Passive & Regulation in 40 CharactersDocument4 pagesCell Membrane Transport: Active, Passive & Regulation in 40 CharactersMahmoud AbouelsoudPas encore d'évaluation
- Cell PhysiologyDocument36 pagesCell PhysiologyRayonesh RayanaPas encore d'évaluation
- Topic 1.4 Membrane TransportDocument8 pagesTopic 1.4 Membrane Transportjasmine wibawaPas encore d'évaluation
- Active TransportDocument7 pagesActive Transportidokofavour2015Pas encore d'évaluation
- Ion transporter: ATP-powered pumps move ionsDocument5 pagesIon transporter: ATP-powered pumps move ionsHafiz AhmadPas encore d'évaluation
- 116 - Transport Mechanisms in CellsDocument4 pages116 - Transport Mechanisms in Cellslastjoe71100% (1)
- 11.05 - Renal Tubules - Mechanisms of ReabsorptionDocument1 page11.05 - Renal Tubules - Mechanisms of ReabsorptionNahin Bin KaysarPas encore d'évaluation
- Membrane-Based Energy Harvest in Mitochondrion Chloroplast With Notes v4-2022-2023Document25 pagesMembrane-Based Energy Harvest in Mitochondrion Chloroplast With Notes v4-2022-2023MARIA ISABILLE DUALLO MUSONGPas encore d'évaluation
- Membrane Transport SystemsDocument33 pagesMembrane Transport SystemsObiajulu Crystal OnyinyechiPas encore d'évaluation
- Transport of Systems in Microbial CellsDocument25 pagesTransport of Systems in Microbial CellsDiptomoy Bhattacharya100% (1)
- Ion Channel: 2 Biological RoleDocument8 pagesIon Channel: 2 Biological RoleMartin MayerPas encore d'évaluation
- Chapter 1 Doc 4Document4 pagesChapter 1 Doc 4Enju ChipepePas encore d'évaluation
- Cell TransportDocument18 pagesCell TransportCallista FeliciaPas encore d'évaluation
- 13-Movement of Substances Across The PDocument8 pages13-Movement of Substances Across The PAbbas TalibPas encore d'évaluation
- Membrane Transport Mechanisms and HomeostasisDocument29 pagesMembrane Transport Mechanisms and Homeostasisramadan100% (1)
- Chapter 11: Membrane Transport: Know The TerminologyDocument25 pagesChapter 11: Membrane Transport: Know The TerminologyShahriar ShamimPas encore d'évaluation
- Bioenergetics: A Novel Theory for Atp SynthesisD'EverandBioenergetics: A Novel Theory for Atp SynthesisPas encore d'évaluation
- GGDocument1 pageGGAnggi AviandriPas encore d'évaluation
- Full House LirikDocument8 pagesFull House LirikAnggi AviandriPas encore d'évaluation
- "The Spirit of Taekwondo": Ukrida Festival Taekwondo Cup 1Document1 page"The Spirit of Taekwondo": Ukrida Festival Taekwondo Cup 1Anggi AviandriPas encore d'évaluation
- What Is Reverse OsmosisDocument7 pagesWhat Is Reverse OsmosisAnggi AviandriPas encore d'évaluation
- AnecdoteDocument1 pageAnecdoteAnggi AviandriPas encore d'évaluation
- Cheat Codes of GTA San AndreasDocument6 pagesCheat Codes of GTA San AndreasAnggi AviandriPas encore d'évaluation
- Unit Unit1Document3 pagesUnit Unit1Anggi AviandriPas encore d'évaluation
- "The Spirit of Taekwondo": Ukrida Festival Taekwondo Cup 1Document1 page"The Spirit of Taekwondo": Ukrida Festival Taekwondo Cup 1Anggi AviandriPas encore d'évaluation
- Association of Sleep Duration With Type 2 Diabetes JurnalDocument18 pagesAssociation of Sleep Duration With Type 2 Diabetes JurnalAnggi AviandriPas encore d'évaluation
- Sleep and Dm2Document7 pagesSleep and Dm2Anggi AviandriPas encore d'évaluation
- Vena, Arteri, DLLDocument5 pagesVena, Arteri, DLLAnggi AviandriPas encore d'évaluation
- Biofarm ADocument6 pagesBiofarm AAnggi AviandriPas encore d'évaluation
- False Positive Malaria Rapid Nnndiagnostic Test in Returning Traveler With Typoid FeverDocument4 pagesFalse Positive Malaria Rapid Nnndiagnostic Test in Returning Traveler With Typoid FeverAnggi AviandriPas encore d'évaluation
- Association between sleep duration and diabetes riskDocument7 pagesAssociation between sleep duration and diabetes riskAnggi AviandriPas encore d'évaluation
- Attract Love CoverDocument1 pageAttract Love CoverAnggi AviandriPas encore d'évaluation
- Bombardier CRJ200 Regional JetDocument2 pagesBombardier CRJ200 Regional JetAnggi AviandriPas encore d'évaluation
- AnecdoteDocument1 pageAnecdoteAnggi AviandriPas encore d'évaluation
- Video Review Jury Sheet PopnasDocument1 pageVideo Review Jury Sheet PopnasAnggi AviandriPas encore d'évaluation
- Pregnancy and Chronic Kidney DiseaseDocument12 pagesPregnancy and Chronic Kidney DiseaseAnggi AviandriPas encore d'évaluation
- "The Spirit of Taekwondo": Ukrida Festival Taekwondo Cup 1Document1 page"The Spirit of Taekwondo": Ukrida Festival Taekwondo Cup 1Anggi AviandriPas encore d'évaluation
- Biol 130 Notes 2012Document121 pagesBiol 130 Notes 2012Nick O'HaraPas encore d'évaluation
- (English) The Nervous System, Part 2 - Action! Potential! - Crash Course A&P #9 (DownSub - Com)Document9 pages(English) The Nervous System, Part 2 - Action! Potential! - Crash Course A&P #9 (DownSub - Com)bank townPas encore d'évaluation
- Foundations of Neuroscience 1632873541. - PrintDocument449 pagesFoundations of Neuroscience 1632873541. - PrintNhật Nam HồPas encore d'évaluation
- ACTIVE TRANSPORT MECHANISMS MOVE SUBSTANCES AGAINST GRADIENTSDocument10 pagesACTIVE TRANSPORT MECHANISMS MOVE SUBSTANCES AGAINST GRADIENTSSylvia DuzonPas encore d'évaluation
- ATP SynthesisDocument5 pagesATP SynthesisHemant SinghPas encore d'évaluation
- Photosynthesis in Higher PlantsDocument17 pagesPhotosynthesis in Higher PlantsJK JHA100% (1)
- Chapter 7 Membrane Structure and FunctionDocument7 pagesChapter 7 Membrane Structure and FunctionIrish Jem BantolinoPas encore d'évaluation
- M and R ModuleDocument247 pagesM and R ModuleMuhammed SamiPas encore d'évaluation
- Mitochondria: The Powerhouses of the CellDocument38 pagesMitochondria: The Powerhouses of the CellPatel BansariPas encore d'évaluation
- Electron Transport and Oxidative Phosphorylation: Paul D. Adams - University of ArkansasDocument43 pagesElectron Transport and Oxidative Phosphorylation: Paul D. Adams - University of ArkansasKrishly Ann Medina SalazarPas encore d'évaluation
- 0 - Final Exam in MajorDocument6 pages0 - Final Exam in MajorKhristine Khate Odiaman MendezPas encore d'évaluation
- A2 Biology Notes - PP & MS - Energy and Respiration - 2019Document226 pagesA2 Biology Notes - PP & MS - Energy and Respiration - 2019Kulsoom JawedPas encore d'évaluation
- Metabolism: Anabolism and CatabolismDocument4 pagesMetabolism: Anabolism and CatabolismMedi OmicPas encore d'évaluation
- Adenosine Triphosphate - WikipediaDocument65 pagesAdenosine Triphosphate - WikipediaBashiir NuurPas encore d'évaluation
- 1 Oxidative PhosphorylationDocument10 pages1 Oxidative PhosphorylationRoland ToroPas encore d'évaluation
- EssentialDocument34 pagesEssentialAdolfo López100% (1)
- Oxidative Phosphorylation and Mitochondrial Physiology - A Critical Review of Chemiosmotic Theory and Reinterpretation by The Association-Induction Hypothesis (Ling)Document68 pagesOxidative Phosphorylation and Mitochondrial Physiology - A Critical Review of Chemiosmotic Theory and Reinterpretation by The Association-Induction Hypothesis (Ling)roan2Pas encore d'évaluation
- ATP Synthesis - BiophysicsDocument13 pagesATP Synthesis - Biophysicsxcjskqt7kwPas encore d'évaluation
- PhotosynthesisDocument281 pagesPhotosynthesisKrishna Priyam SinghPas encore d'évaluation
- M11 Active Transport M11Document15 pagesM11 Active Transport M11Samantha Kyle E. PedrigozaPas encore d'évaluation
- Mitochondria and Chloroplasts MCQsDocument14 pagesMitochondria and Chloroplasts MCQsJenny SmithPas encore d'évaluation
- Chapter 12 & 13: Energy, respiration and photosynthesisDocument88 pagesChapter 12 & 13: Energy, respiration and photosynthesiseric sivaneshPas encore d'évaluation
- ChemiosmosisDocument7 pagesChemiosmosisSkenzKenzPas encore d'évaluation
- PBIO211Lec FINALS 1Document26 pagesPBIO211Lec FINALS 1richard respetoPas encore d'évaluation
- PoB I - CA 1 - Topic 6 - Wrestling With Weight LossDocument4 pagesPoB I - CA 1 - Topic 6 - Wrestling With Weight LossGibran Rifa'iPas encore d'évaluation
- Mechanisms of Energy Transduction by Charge Translocating Membrane ProteinsDocument41 pagesMechanisms of Energy Transduction by Charge Translocating Membrane ProteinsVatra ReksaPas encore d'évaluation
- Advanced Physio Final Exam PracticeDocument111 pagesAdvanced Physio Final Exam PracticefoufouPas encore d'évaluation
- Bİochem 5Document79 pagesBİochem 5john doePas encore d'évaluation
- Parte 03Document100 pagesParte 03Pâmella PicançoPas encore d'évaluation
- Kazakh National Medical University Course on Biological MembranesDocument78 pagesKazakh National Medical University Course on Biological MembranesSyed Inam UllahPas encore d'évaluation